
АМИНОКИСЛОТЫ В МЕХАНИЗМАХ АДАПТАЦИИ В ЭКСПЕРИМЕНТЕ И У СПОРТСМЕНОВ В РАЗНЫХ ВИДАХ СПОРТА
Павлов В.А. и другие
АННОТАЦИЯ
В исследовании уточняется значение свободных аминокислот в регуляции адаптивного ответа на стрессовые воздействия. В эксперименте на животных (морские свинки и крысы) установлено принципиальное различие их защитных адаптивных механизмов по отношению к таким повреждающим факторам, как и токсическое воздействие ксенобиотиков, мутагенные ксенобиотики, заражение микобактериями туберкулеза, кровопотеря. Эти различия во многом определяются содержанием в тканях животных аминокислот-адаптогенов и их производных. Таких как таурин, глутатион, глутамат, цистеин, глицин и ряд других.
В проведенных далее исследованиях на спортсменах разных видов спорта (легкоатлеты, борцы, хоккеисты) установлено, что аминокислоты-адаптогены и у человека при интенсивных спортивных стрессовых нагрузках во многом определяют возможность эффективной адаптации к ним и соответственно успех тренировок. Так для легкоатлета «стержневыми» аминокислотами успеха являются аспартат и глутамин, для борцов-глутамат, для хоккеистов-таурин. Эти отличия определяют различия адаптивного ответа на хронические стрессовые спортивные нагрузки.
Книга может представлять интерес для физиологов, биохимиков и специалистов, применяющих аминокислоты в спорте и фитнесе.
Рецензенты:
Котомцев Вячеслав Владимирович, доктор биологических наук, профессор, старший научный сотрудник Института иммунологии УрО РАН
Вашляев Борис Федорович мастер международного класса по конькобежному спорту, кандидат педагогоческих наук, профессор кафедры теории и методикифизической культуры УрГУФК.
ВВЕДЕНИЕ
Достижения современной науки в области биологии граничат с фантастикой. Опыты над генетическим материалом начиная от микроорганизмов до человека дают многообещающие результаты. Трансплантация органов, тканей, культуры клеток произвели революцию в ряде отраслей медицины. Генетики замахиваются на искусственное создание новых организмов, улучшение генетического материала человека и другие казавшиеся невероятными ранее биотехнологии. Предпринимаются попытки довести до совершенства адаптивные возможности организма, например, в большом спорте, за счет применения различных методик тренировки, биологически активных добавок, фармакологических препаратов.
Однако давно известно, что каждый организм индивидуален и как раз от этого во многом зависит успех всех применяемых биотехнологий. От чего же зависит эта индивидуальность?
Имеются достаточно убедительные результаты исследований, что главенствующую роль в строении и регуляции живой материи имеют аминокислоты, соотношение которых создает среду, контекст на котором разворачиваются эволюционные процессы ее совершенствования. Ранее мы установили, что в реализации адаптивных механизмов животных и человека могут иметь регуляторные аминокислоты-адаптогены образующиеся преимущественно в мозге или печени. Роли свободных аминокислот в совершенствовании адаптивных механизмов организма в эксперименте на животных и спортсменах – профессионалах и посвящено это исследование.

1. РОЛЬ АМИНОКИСЛОТ В МЕХАНИЗМАХ АДАПТАЦИИ К СТРЕССОВЫМ НАГРУЗКАМ
1.1 АМИНОКИСЛОТЫ В МЕТАБОЛИЧЕСКОЙ «РЕКЕ» ФИЛОГЕНЕЗА
На заре появления живой материи при высокой температуре, давлении, радиации и других жестких условиях появились первичные органические молекулы: углеводы, жиры, аминокислоты. Конденсация этих веществ привела к появлению первичной протоплазмы с первичным метаболизмом, протекание которого целиком зависело от состояния окружающей среды, параметры которой и обеспечивали необходимую энергию протекания метаболических процессов в коацерватах (первичных функциональных единицах живой материи) а затем в первичных одноклеточных организмах (Опарин Л.И. «Возникновение жизни на земле» 1958 г.).
Протекание метаболических реакций при снижении температуры, давления, радиации и других внешних воздействий было возможно лишь при появлении белков-ферментов, резко понижающих энергетический порог этих реакций, т.е. энергия жестких внешних воздействий аккумулировалась в энергии полипептидной цепочки аминокислот, взаимодействующих друг с другом в виде полипептидных связей и составляющих полипептиды и белки. Ферменты и другие структурно-функциональные единицы живой материи включают в себя элементы неорганических веществ, то есть, неживой материи — железо, медь, кобальт, никель и др., входящих в активные центры белков — ферментов.
Моделирование условий, в которых согласно теории Опарина-Холдейна зарождалась жизнь на земле, приводит к абиогенному синтезу 4-х аминокислот, входящих в состав белков всех организмов — это глицин, аланин, глутаминовая и аспарагиновая аминокислоты — «древние аминокислоты». Эти аминокислоты и являются родоначальниками других аминокислот. Вступая во взаимодействие с соединениями серы, ароматическими веществами, претерпевая молекулярные перестройки, образуется все разнообразие известных на сегодня аминокислот. Аминокислоты выполняют выдающуюся роль в реализации метаболизма и функционирования органов и систем организма. Многие из их производных (ГАМК, ГОМК и тд.) являются адаптогенами, т.е. веществами, выполняющими защитные функции или принимающие участие в регуляции защитно-адаптивных систем организма. Суть их заключается в антиоксидантном потенциале, защите от воздействия свободных радикалов, последствий воздействия радиации, в процессах обезвреживания токсичных веществ. Это энергетический и пластический ресурс организма. Они стабилизируют водно-электролитный баланс, клеточные мембраны и их ионные насосы [22,24,158,159,352,357,197,198,200,201,202,34,35,36,157,576].
Неся на себе положительный и отрицательный заряд, а также, обладая кислотными и основными свойствами, они поддерживают электролитный баланс, формируют буферные системы, рН, а так же другие важные гомеостатические системы клеток и тканей. Необходимо отметить, что в состав белков многоклеточных организмов входят только L изомеры аминокислот, тогда как микробные клетки могут содержать D изомеры. С участием аминокислот (аргинина, метионина, глутамина и др.) происходит формирование систем дифференцировки и защиты организмов [378].
По мере продвижения по филогенетической лестнице у организмов происходит упорядочение набора аминокислот и включение их в алгоритм реализации адаптивных механизмов. Так, у беспозвоночных животных фонд аминокислот чрезвычайно разнообразен и малоизучен. В тканях насекомых встречается огромное количество аргинина, пролина, глицина и глутаминовой кислоты, а так же амидов — глутамина и аспарагина, которые могут составлять более половины фонда свободным аминокислот. Целесообразность такого набора аминокислот у насекомых до сих пор остается не ясной. То же самое относится к таурину, глицину у морских моллюсков. (Хочачка П., Сомеро Дж. 1977, 1988) [357]
У высших животных содержание аминокислот приобретает тканевую и органную специфичность, строго постоянную для каждого вида. Кровь является связующим звеном между органами и системами в плане обеспечения и перераспределения аминокислот. Непрерывно протекающие процессы анаболизма и катаболизма белков определяют положительный или отрицательный азотистый баланс, характерный для процессов роста, адаптивных процессов при стрессовых воздействиях и патологических состояниях. В норме у взрослого организма наблюдается устойчивое равновесие аминокислот [2,92].
Известно, что отдельные аминокислоты при определенных условиях сами собираются (спонтанно) в полипептидные цепочки, которые затем скручиваются во вторичные и третичные структуры — способные уже проявлять свойства белков [34,36,42,43,235].
Из данных литературы и наших собственных работ следует, что аминокислоты, помимо того, что они являются кирпичиками построения белков, биологически активных веществ в организме и других его важнейших структур, выполняют важнейшую функцию создания метаболического (обменного) фона, который определяет адаптивные возможности организма и их регуляцию. Т.е. соотношения аминокислот и их биологически активных производных в тканях организма являются своеобразной математикой живой материи, от изменений пропорций которой зависят защитно-адаптивные механизмы, регулируемые соотношением аминокислот [236,237,238,239].
Различные комбинации этих метаболитов могут создавать огромное количество вариантов (миллиарды) метаболических типов, создавая специфику обмена веществ у того или иного организма, а также адаптивные и метаболические особенности различных видов. В то же время количество нуклеотидов, из которых строятся нуклеиновые кислоты (ДНК и РНК), лежащие в основе хромосомного аппарата клетки и его воспроизводства, составляет всего 4 основных и два добавочных, из которых может образоваться 64 варианта кодонов, то есть основных элементов, которыми записывается генетический код. Учитывая приведенные обстоятельства и наши знания о воспроизводстве наследственной информации, как и филогенетической информации вообще, следует признать, что они еще очень недостаточны, для того, чтобы пытаться искусственным или полуискусственным путем без непредвиденных отрицательных последствий создавать новые организмы (клонирование, трансгенный перенос, попытки найти гены, ответственные за те или иные заболевания, и излечить эти заболевания путем отключения или стимуляции участков хромосом и т.д.). Появляются весьма аргументированные точки зрения, что генетика в современном ее состоянии вообще является лженаукой, и истинное ее развитие является делом будущего, при достижении человечеством более высоких научных и мировозренческих горизонтов [91,142,501]
Хорошо известен факт, что содержание свободных аминокислот в тканях обладает высоким постоянством и специфичностью, то есть для каждого вида животного, или даже отдельной особи, оно строго постоянно в его тканях [235]. Изменение количества аминокислот в тканях сопряжено со значительными перестройками защитно-адаптивных механизмов и выраженные отклонения наблюдаются при тяжелых патологических процессах (различных заболеваниях, травмах, старении и т.д.). Сопоставление содержания свободных аминокислот в тканях различных животных говорит о том, что у животных, находящихся на более ранних ступенях филогенетического развития общее содержание аминокислот в тканях больше, чем у животных, находящихся на более высоком уровне развития.
В то же время качественный состав, разнообразие биологически активных веществ-производных аминокислот и ряда самих аминокислот, обладающих такой активностью, возрастает по мере филогенетического совершенствования организмов (метаболическая река филогенеза) [357,158,244,126]. Особенно наглядно это видно по изменениям количества таурина и глутаминовой кислоты. Так, содержание таурина наиболее значительно в тканях организмов, стоящих на более древних, примитивных филогенетических уровнях. Затем по мере филогенетического совершенствования и появления высокоорганизованных защитно-адаптивных систем, количество таурина в тканях постепенно снижается, а глютаминовой и аспарагиновой аминокислот, триптофана, фенилаланина и их производных — увеличивается [126, 158,262, 263, 264, 365]
И самое большое количество таких биологически активных веществ наблюдается в тканях приматов и человека, особенно в их головном мозг [22]. Установлено, что стрессовые воздействия сопровождаются повышенным образованием метаболитов-адаптогенов в нейроэндокринной системе. В частности, в нейронах коры головного мозга образуются такие вещества как нейропептиды (в настоящее время известно более 600 биологически активных нейропептидов), эндорфины, энкефалины, производные ряда аминокислот (глютаминовой, аспарагиновой, триптофана, фенилаланина, гистамина), являющиеся гормонами любви, счастья, ненависти, страха, воли и других проявлений духа, имеющего воплощение в организме, материи, обществе [22,155,319-328,320,352,360].
Возможно, именно так и шел процесс адаптации первичной живой материи к изменяющимся условиям на планете, при использовании колоссального модификационного ресурса аминокислот. В этом плане следует отметить, что нуклеотиды — основные компоненты нуклеиновых кислот хранителей наследственной информации является по сути дела производными нескольких аминокислот: глицина, аспарагиновой кислоты, глутамина [35,235,291,155,156]. А важнейшие регуляторы биосинтеза нуклеиновых кислот и пролиферативных процессов вообще — полиамины производные глутамата (орнитина, аргинина), метионина, лизина. Другие важнейшие регуляторы метаболизма: катехоламины, гормоны щитовидной железы (тироксин, тиронин), гистамин и серотонин, глутатион и карнозин (анзерин), а также многие другие биологически-активные вещества являются производными аминокислот [158]. Таким образом, важнейшими регуляторными метаболитами защитно — адаптивных систем организма являются либо сами аминокислоты, либо их производные.
Поэтому вполне возможно, что свойства первичного метаболического субстрата во многом определялись особенностями аминокислот, входящих в него, а при изменении внешних факторов изменялись и эти особенности в сторону превращения аминокислот в регуляторные метаболиты. Изменение соотношения аминокислот-адаптогенов и их биологически активных производных, видимо лежало в основе совершенствования адаптивных механизмов.
Все это могло резко повышать приспосабливаемость органической материи к меняющимся условиям среды. И вершиной этого процесса стало появление из азотистых метаболитов нуклеиновых кислот и хромосомного аппарата. Все это предопределило формирование клеток и координированного воспроизводства филогенетической информации. Однако не следует переоценивать значение нуклеиновых кислот и хромосомного аппарата — это лишь верхняя ступень регуляторной пирамиды метаболического гомеостаза — средние и нижние ступени в ее памяти сохранились и заняты они белками (полипептидами), олигопептидами и аминокислотами. И вся эта информация воспроизводится в процессе полового и бесполого размножения, составляя метаболическую реку, в которой ключевое значение имеют взаимоотношения и особенности обмена важнейших аминокислот-адаптогенов.
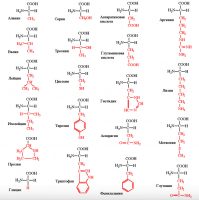
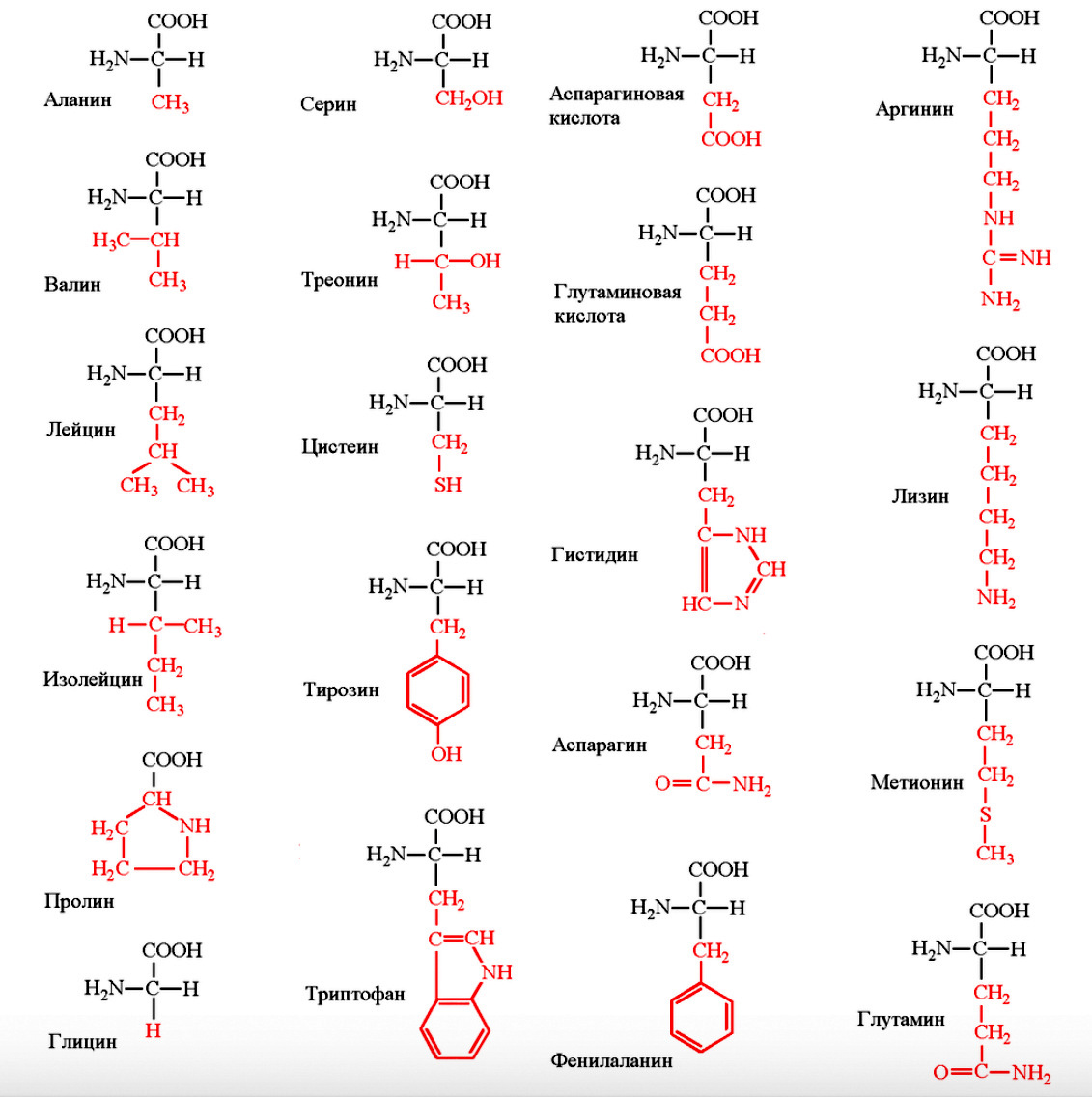
Приводим строение 20 главных аминокислот необходимых для построения белков и биологически активных регуляторных метаболитов (рис 1.)
Рис.1 Строение и название аминокислот:
В настоящее время насчитывается более 200 различных производных основных аминокислот, что в самых различных комбинациях может нам дать колоссальное количество информации, из которой наиболее ценная, в той или иной стресс-ситуации, отбирается на уровне фенотипа и только затем фиксируется на уровне генотипа. Это можно подтвердить теми фактами, когда адаптивный ответ включается без структурного следа адаптации.
Имеются различные свидетельства того, что наследственная информация содержится и передается не только через гены, входящие в нуклеиновые кислоты. Наиболее известный пример: телегония.
Наследственная информация передается не только через хромосомы, а, возможно, через тот метаболический фон, который во многом может определяться спектром аминокислот — регуляторов-адаптогенов.
Аминокислоты в этом информационном потоке могут выполнять роль неспецифического базисного метаболического фона различных уровней филогенетической лестницы. Адаптивные механизмы, начиная с примитивных форм жизни, базируются на обмене аминокислот и их соотношениях, определяющих заряды и структурные элементы протоплазмы, получение энергии, стабилизации водно-солевого обмена, рН и других важнейших параметров функции клетки [357,235,170].
Наибольшее регуляторное значение имеют аминокислоты-адаптогены, обогащенные азотом (глутаминовая, глутамин, аргинин и ряд других) или имеющие в своем составе серу (метионин, цистеин, таурин и некоторые другие). Важнейшее регуляторное значение имеют производные аминокислот – олиго- и полипептиды: полиамины, гаммаглутамилпептиды, имидазолпептиды и ряд других [158,357,24,431,562,576].
Это значение является следствием того, что азот и сера имеют переменную валентность и могут подвергаться различной степени окисления, то есть менять свои химические свойства. Азот имеет на своих энергетических уровнях 7 электронов, причем на последнем — 4 электрона; сера имеет на своих энергетических уровнях 16 электронов и на последнем 6 электронов. Из чего следует, что сера имеет больший диапазон окислительно-восстановительных превращений и поэтому неслучайно, что в процессе эволюции природа выбрала именно серу, как важнейший инструмент в регуляции механизмов обмена веществ в живой природе. Так, азот может окисляться от 0 валентности (N2 азот воздуха) до +5, например, в составе аммиака. Сера окисляется от 0 валентности (ископаемая сера) до +6 в сернистом ангидриде или даже до -2 в сероводороде. Отмеченный диапазон окислений, определяет образование различных метаболитов, которые участвуют в регуляции различных метаболических процессов. Так, например, восстановленная сера (SH) является ключевым метаболитом для поддержания нормального функционирования многих ферментов организма, а таурин (содержащий SО3) является мощным антиоксидантом и стабилизатором мембран; остаток серной кислоты (SО4) входит в кислые гликозаминогликаны соединительной ткани. Окисленная сера входит во многие структурные компоненты клеток, например, она необходимая для образования соединительной ткани — гепарин, хондроитин-4-сульфат, хондроитин-6-сульфат, то есть, кислых гликозамингликанов. Имеются сведения, что соотношения кислых и нейтральных гликозаминогликанов определяют различия метаболизма и функциональных состояний важнейших адаптивных систем организма (кроветворная ткань, лимфоидная ткань)[385].
Большое значение имеет количество и состояние окисленности железа и серы в активных центрах белков ферментов (Fe-S) [361]. В свою очередь, повышение в тканях метаболитов, обогащенных азотом (глутамин, полиамины, нуклеиновые кислоты) отчетливо стимулируют пролиферативные процессы. Поэтому соотношение содержания серы в тканях и степени ее окисленности с одной стороны и азота с другой стороны являются важнейшим регуляторным механизмом в направленности метаболических адаптивных особенностей организма — N и S тип. Но сера и азот в тканях представлены в соответствующих серо и азотсодержащих аминокитслотах. Таким образом, из имеющихся данных следует, что в адаптивных механизмах преобладают N или S аминокислоты.
Совершенствование форм жизни сопряжено с изменениями азотистого обмена и ресурсов аминокислот — переход от аммониотелии к урикотелии и уреотелии [357].
Исходя из концепции пифагорейцев, окружающая нас природа выражена в определенных соотношениях, достигающих определенного уровня гармонии, в наибольшей степени это проявляется в живых организмах, существующих вопреки неблагоприятным условиям окружающей среды, благодаря высокой степени организации живой материи. Наивысшей степени совершенства достигают аэробные организмы, прежде всего человек, использующие для своей жизнедеятельности окисление поступающих с пищей веществ кислородом. Однако это же обстоятельство определяет повреждающее его воздействие в условиях стрессовых нагрузок, особенно превышающих адаптивные ресурсы организма.
Кислород и его активные радикалы могут повреждать все ткани организма, особенно его многофункциональных систем: нейроэндокринной, дыхательной, сердечно-сосудистой, выделительной и других, что и лежит в основе патогенетических нарушений при многих патологических состояниях, в том числе и при старении. Защитными свойствами по отношению к агрессивным радикалам кислорода обладает сложно устроенная антиоксидантная система. По сути, весь организм можно рассматривать как высокоорганизованную антиоксидантную систему, предназначенную для сохранения информации, содержащейся в половых клетках. Исходя их этих представлений защитно-адаптивныe возможности организма, его здоровье и продолжительность жизни во многом зависят от состояния его антиоксидантных систем, обеспечивающихся, в свою очередь, необходимыми ресурсами N и S аминокислот.
У млекопитающих все регуляторные метаболиты либо сами являются аминокислотами, олигопептидами, полипептидами, либо производными аминокислот. Все это создает некий метаболический порог стресса, выход, за который означает развитие дистрессового состояния или патологию. Накапливается все больше фактов о том, что возвращению к физиологическому регуляторному уровню (структурный след адаптации) можно способствовать введением в организм, который подвергается какому-либо запредельному воздействию, ряда аминокислот-адаптогенов (глутамата, различные полиптиды и смеси аминокислот, полиаргинина, цистеина, метионина, олигопептидов — тафтсин, тимоген) [117, 221, 223, 224, 228, 332,237, 238, 239, 400, 404, 407, 408, 410, 414, 417, 418, 557].
Соматические клетки могут делиться 50-70 раз, в то время как половые клетки могут делиться бесконечно, то есть обладают практически бессмертием, их существование поддерживается их окружением или всем организмом, в которых эти половые клетки образуются. Так, организмы выполняют защитно-адаптационные функции по поддержанию метаболического потока сконцентрированного в половых клетках. При слиянии мужских и женских половых клеток происходит метаболический взрыв, сопровождающийся расширением метаболической информации, ибо помимо слияния ядерного аппарата половых клеток происходит слияние метаболитов, вводимых со спермой с метаболитами, окружающими яйцеклетку. В женском организме после оплодотворения происходят радикальные изменения, направленные не вынашивание плода, а затем и на успешное родоразрешение.
Расширение это доходит до стадии половозрелого состояния организма, а затем начинает постепенно уменьшаться вплоть до полной утраты его адаптивных возможностей. На смену исчерпавшему свои защитно-адаптивные возможности приходит другой организм.
Диапазон возможностей организма широк в начале жизни. При этом количественно возможности адаптации к каждому конкретному условию расширяются до определенного возраста. С возрастом список условий, к которым адаптируется организм ограничивается наличными условиями, условия же, которые не оказывают влияния на организм не требуют к себе адаптации и возможности адаптации организма к этим отсутствующим условиям с возрастом снижаются. К старости диапазон возможностей организма сужается до строго определенных, что и является предпосылкой к смерти.
С этих позиций процесс зачатия новой жизни в материнском организме, это не есть только слияние собственно наследственного материала, несущего память о филогенезе, но и слияние двух метаболизмов, несущих информацию о двух актуальных онтогенезах. Стресс-воздействия могут существенно менять оперативный уровень метаболизма или фенотип того или иного индивидуума, но на генетическом уровне эта информация откладывается лишь в случае адекватности воздействий метаболическому уровню той или иной ступени филогенетической лестницы.
На ранних стадиях онтогенеза плод пользуется метаболическим фондом матери, на который накладывается вводимый с семенем метаболический фонд отца [362].
По мере созревания плода меняются пропорции метаболического фонда, что предопределяет основные параметры роста и развития организма. По мере взросления осуществляется некий баланс в системе N/S. Однако, если на ранних этапах развития преобладает S тип, то по мере старения организма возрастает доля N типа. Метаболическая память организма отличается от генетической своей высокой мобильностью и при неблагоприятных воздействиях, превышающих стрессовый порог, возможен откат на более древние метаболические системы (например, при массивной кровопотере или травматическом шоке начинает преобладать аммониотелический тип азотистого обмена, характерный для беспозвоночных). Это позволяет пережить повреждающее воздействие и затем выйти на физиологический уровень. Введение в организм подопытных животных аминокислот-адаптогенов, например, глутаминовой кислоты, значительно ускорит этот процесс, что препятствует развитию необратимых патологических процессов [40,228, 232, 236 , 233, 263, 264, 458, 459,68,238,509,572,593].
В этом плане, необходимо указать на то, что эксперименты по клонированию животных и методы генного инженеринга не могут быть успешными и безопасными без учета этих обстоятельств, ибо полноценная филогенетическая информация несется не только с генами, но и со всей той средой, которая окружает половые клетки. Так, сперма содержит большое количество полиаминов и богатых азотом аминокислот: аргинина, лизина, таурина, в то время как готовая к оплодотворению слизистая матки содержит большое количество серосодержащих аминокислот, серы и глутатиона и богатых SH группами белков и метаболитов (Алов И.А., Брауде А.И., Аспиз М.Е. «Основы функциональной морфологии клетки» 1966).
В ходе оплодотворения и дальнейшего развития плода меняется метаболический фон как вокруг плода, так и в женском организме вообще. Изменяются пропорции аминокислот с повышенным количеством азота (N — глутамин, аргинин, полиамины и некоторые другие) и серы (S — метионин, цистеин, таурин и ряд других), регуляторных метаболитов N и S типов. После рождения плода изменение этих пропорций продолжается вплоть до достижения половозрелого состояния. Этот период у разных классов и видов является тем периодом, когда организм в результате различных внешних воздействий способен максимально к ним приспосабливаться, т.е. включаться в филетический поток информации, вносить в него свою лепту и передавать далее по наследству [397,441].
С возрастом, испытывая на протяжении жизни повреждающие (стрессовые) воздействия, меняется метаболический ресурс организма. Эти изменения у аэробных организмов, использующих в окислительно-восстановительных процессах (ОВП) кислород, связаны, прежде всего, с тем, что снижаются антиоксидантные ресурсы и антирадикальные ресурсы вообще. Т.е. защитные механизмы, направленные на предохранение клеток, и тканей (прежде всего мембранных структур) от повреждения кислорода и его активных форм (АФК), возникающих в ходе его использования в качестве окислителя в обменных процессах [19, 20, 29, 30, 59, 199, 15, 133, 29, 181, 260, 442, 499, 568, 589, 416].
Таким образом, заложенная в метаболическом фонде программа подразумевает в процессе жизнедеятельности организма постепенное «стирание» этого фонда о стресс — воздействия или точнее повреждающее воздействие 02 и особенно образующиеся в процессе метаболизма его активные формы в меняющихся условиях среды. Изменение метаболического фонда ведет к дисбалансу в системе.
Процессы старения у человека касаются, прежде всего, нервной ткани, особенно коры головного мозга и соединительной ткани, т.е. структур, ответственных за обеспечение метаболических процессов организма аминокислотами, входящими в семейство глутамата и серосодержащими аминокислотами-адаптогенами. При этом происходит прогрессирующее снижение SH групп в регуляторных белках — ферментах и других важнейших адаптивных системах организма, ответственных за их функциональное состояние, с последующими нарушениями в азотистом, жировом, углеводном обмене и развитием тех или иных патологических состояний старения [11, 29, 30, 88, 102, 185, 188, 300, 301, 441, 442, 479].
Диспропорции в ресурсах регуляторных N- и S- аминокислот помимо периода старения характерны и для периода созревания организма. Однако, если в период старения преобладают и прогрессируют N-системы со склонностью к воспалительно-дистрофическим пролиферативным (опухолевым) процессам, то в детском возрасте преобладает S-тип с гиперконтролем над пролиферативными процессами и относительной редкостью дистрофически-опухолевых процессов в этот период [263, 223, 224, 358].
Избыток серосодержащих адаптогенов защищает головной мозг ребенка от гипоксии и родовой травмы [22]. Однако преобладание S-типа делает организм ребенка менее устойчивым к большинству инфекций и токсических воздействий. Такое преобладание, очевидно, обусловлено тем, что голова и мозг ребенка имеют значительно больше размеры по отношению к другим частям тел, чем у взрослого человека. При этом интенсивность метаболических процессов: окислительно-восстановительных и образование серосодержащих аминокислот и глутамата наиболее интенсивны в раннем детском возрасте в головном мозге ребенка с постепенным снижением этих параметров по мере роста и взросления организма [263,264]. А в процессе старения идут обратные процессы с уменьшением объема мозга, интенсивности в нем ОВП, образования глутамата и серосодержащих метаболитов. Болезни, приводящие к снижению ОВП мозга, сопровождается снижением образования в нем глутамата и серосодержащих аминокислот и угнетением адаптивных возможностей организма (болезнь Альцгеймера, Паркинсона, Дауна и др.).
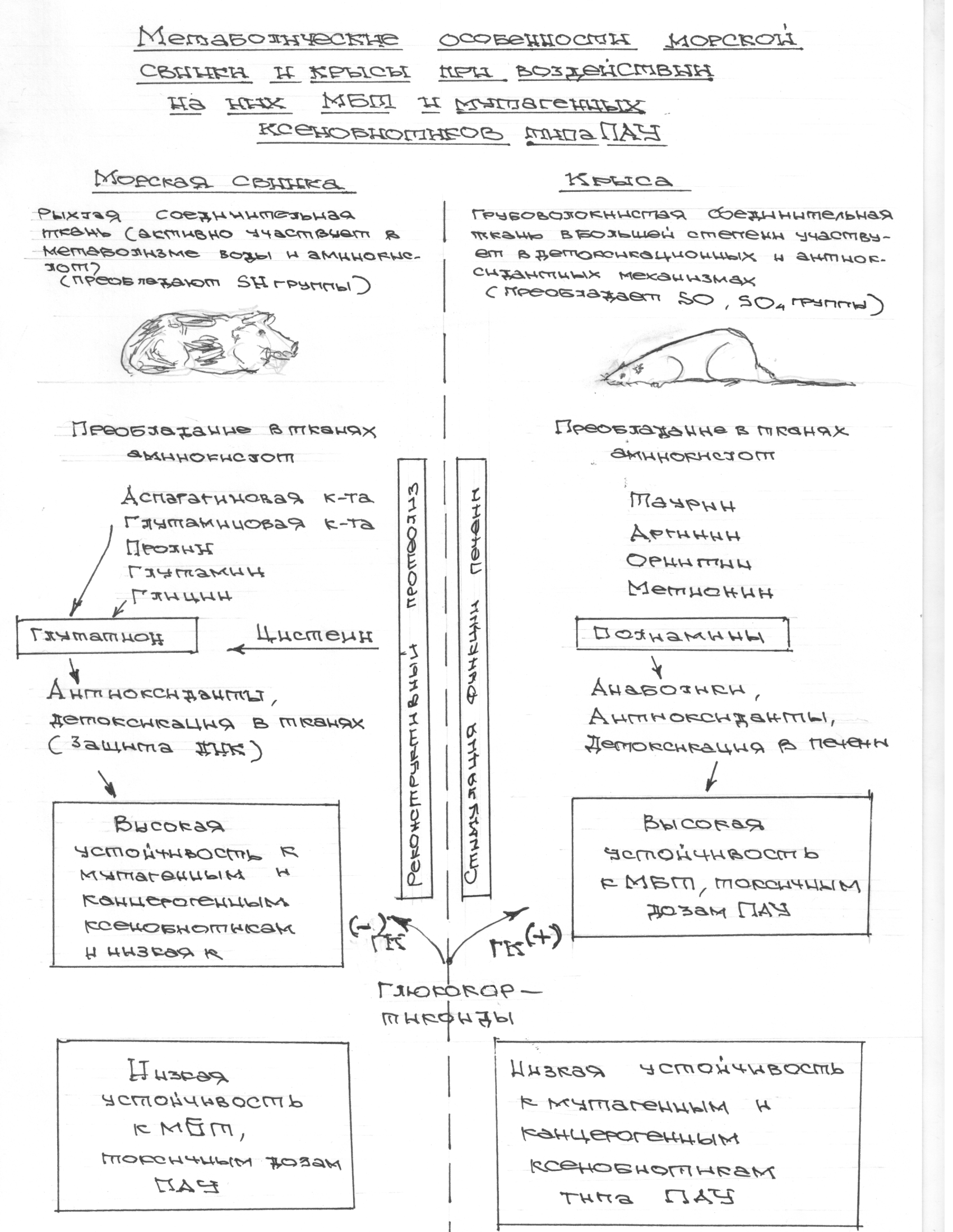
В последние годы в развитых индустриальных странах нарастает проблема изменений у человека состояния соединительной ткани. Все чаще развивается так называемая дисплазия соединительной ткани (ДСТ). Эти нарушения сопровождаются нарушениями соматического здоровья, иммунитета, психики, нарушениями в процессах воспитания, образования, коммуникаций и т.д. Приводятся данные, что до 70% детей имеют ту или иную степень ДСТ и отклонения в психическом развитии (дисгенетический синдром — ДС), сопровождающиеся нарушениями формирования речи, коммуникаций, социализации, требующие больших усилий в коррекции данных нарушений. Если такой своевременной коррекции нет, то развитие и формирование личности происходит с дефектом. Причем дефект это может быть очень большим — вплоть до инвалидизации [296]. На основании наших многолетних исследований на морских свинках и крысах было установлено, что в адаптивных механизмах защиты организма имеют большое значение особенности строения соединительной ткани. [8,296.300,301]
У морских свинок соединительная ткань тонковолокнистая и рыхлая, а у крыс – грубоволокнистая и плотная, что хорошо видно при окрашивании тканей на коллаген. При этом объем соединительной ткани у морских свинок ничуть не меньше, чем у крыс. Соединительная ткань имеет важнейшее значение в механизмах детоксикации и выведения из организма токсичных веществ и одновременно является своеобразным метаболическим ресурсом для нейроэндокриной системы (аминокислоты, углеводы, липиды, витамины, минеральные вещества и т. д.).[259,300,301]
Имея существенную разницу по строению соединительной ткани морские свинки и крысы имеют совершено различную устойчивость ко многим факторам воздействия со стороны внешней среды. Так крысы высокоустойчивы к заражению большинством болезнетворных микробов, токсическому воздействию разных веществ, перепадам температур и другим экстремальным факторам. Морские же свинки высокочувствительны к этим факторам и быстро погибают при их воздействии. Но морские свинки высокоустойчивы к мутагенам и канцерогенам, то есть веществам вызывающим генетические мутации и злокачественные опухоли [236,237,238,239].
По нашим данным этот связано с тем, что морские свинки располагают высокоактивной системой глутатиона (это трипептид, состоящий из трех аминокислот – глутаминовой, глицина и цистеина, и способный интенсивно окисляться и восстанавливаться, имеет решающее значение в защите ядерного аппарата клетки от мутагенов и канцерогенов) подпитываемой ресурсами соединительной ткани. У крыс этого нет и на них легко моделируется как мутагенез, так и канцерогенез, несмотря на их устойчивость к мощным повреждающим факторам за счет их мощной печени и грубоволокнистой соединительной ткани [85, 139, 164, 166, 175, 227, 357, 358, 359].
Такая разница может быть связана с тем, что морские свинки, проживая на островах недалеко от Южной Америки либо получали с пищей избыток аскорбиновой кислоты, либо подвергались повышенному воздействию радиации (глутатион и другие серосодержащие аминокислоты защищают организм от ионизирующего излучения), либо получали с пищей то, чего нет в других районах Земли и что требует больших количеств глутатиона и серосодержащих аминокислот для обезвреживания, либо сочетание всех этих факторов. Но факт остается фактом, морские свинки — единственные экспериментальные животные, не способные синтезировать аскорбиновую кислоту и производят большое количество глутатиона в тканях взамен ее. В этом смысле они близки к человеку. Крысы же являясь своеобразным биологическим реликтом (первые млекопитающие были похожи на крыс) и, проживая в загрязненной среде и подвергаясь разнообразным неблагоприятным воздействиям её, что бы выжить, сохранили способность противостоять многочисленным отрицательным факторам, и, если нужно, быстро мутировать. Но при этом они вынуждены пожертвовать устойчивостью к мутагенам ядерного аппарата. Поэтому у них и не выражена глутатионовая защита.
Во внутриутробном периоде жизни мозг плода вместе с матерью принимает участие в регуляции собственного метаболизма, причем те же химические вещества необходимы для реализации такого взаимодействия (таурин, цистеиновая кислота, глутатион и др.). По мере роста и развития ребенка, роль метаболических и интуитивных механизмов адаптации с участием мозга утрачиваются, равно как утрачиваются ненужные на определенном этапе онтогенеза структуры. Роль мозга как поставщика метаболитов резко снижается и возрастает его роль как нервного управляющего центра в первые годы жизни ребенка [263,264,22]. Одновременно меняется и роль соединительной ткани – из метаболического ресурса мозга она превращается в самостоятельную структуру с собственными функциями. При этом в метаболизме все большее значение начинает приобретать печень (липопротеиды, иммунные белки, системы детоксикации еще много чего свойственное печени как «биохимической лаборатории» организма), координирующая функции важнейших адаптивных систем организма [78,43,175,235,232,263,264].
Человек становится взрослым, способным переносить различные стрессовые воздействия окружающей среды. Для наиболее полной реализации возможностей человека необходимо, чтобы внешняя среда для детского организма СОЗДАВАЛА УСЛОВИЯ ОПТИМАЛЬНОГО РАЗВИТИЯ ИМЕННО ФОРМИРУЮЩИХСЯ В ДАННЫЙ МОМЕНТ СТРУКТУР МОЗГА. Но в современных условиях, с культом естественного отбора, образование превратилось в своеобразный спорт – у кого ребенок раньше начнет читать, писать и считать.
Уже не редкость двух-трехлетние грамотные детки, и остановить родительский ажиотаж в этом направлении не представляется возможным. Государственные структуры, несущие ответственность за будущее, поощряют этот перекос – начало школьного обучения сместилось с 8-9 лет в 1900 году, до 6 лет к 2000 году, и речь уже идет о том, чтобы посадить за парты пятилетних в детском саду. У ребенка мозг еще не научился управлять телом, а перед ним уже ставится задача овладения письменной речью. Разрушение семьи и утрата общинного воспитания нарушили развитие структур мозга ответственных за коммуникации. Ребенок, слабо адаптированный к коммуникациям с себе подобными не видит смысла в кооперации, как следствие в речи он НЕ СТРЕМИТСЯ БЫТЬ МАКСИМАЛЬНО ПОНЯТЫМ, его речь все больше становится рудиментом, а центры речи, ранее обеспечивавшие социализацию ныне все больше подчинены древним отделам мозга, работающим в интересах гомеостаза.
И по факту мы видим, что МЕТАБОЛИЧЕСКИЕ АДАПТИВНЫЕ ВОЗМОЖНОСТИ СОХРАНЯЮТСЯ ОЧЕНЬ ДОЛГО, и, как следствие, печеночные возможности метаболизма не формируются в полной мере. Поэтому соединительная ткань, как ресурс нейроэндокринной системы вынуждена сохранятся в мобильном рыхлом состоянии, что и приводит к ДСТ со всеми вытекающими последствиями.
Имеются данные, что в популяции людей, имеющих «многовековой городской стаж» (как и у других «городских обитателей») в мозге исчезают, либо морфологически резко изменены структуры, ответственные как за движения (красное ядро), так и за коммуникации (высшая кора, мозолистое тело). ДСТ и Дисгенетический Синдром (ДС) ведут к индивидуальной и социальной психопатии с агрессивным навязыванием своего мировоззрения всем окружающим при одновременной неспособности радикально решать все более нарастающие перед обществом проблемы в силу утраты физиологической и психологической адаптивности) [296].
Для человека, как вида, это путь в вырождение. Но на планете много народов, сохранивших нормальную природную адаптивность именно они должны взять на себя лидирующую роль в развитии человеческой культуры. По данным наших исследований на спортсменах, здоровых людях, не занимающихся спортом, до 50% составляют люди в адаптивных механизмах которых преобладает печень.
Но, как мы уже отмечали выше, психологи говорят о том, что на смену им идет городское детское население с 70-80% ДСТ и ДС [296]. Так что ситуация требует безотлагательного изменения систем воспитания, образования и образа жизни наших детей.
1.2 АМИНОКИСЛОТЫ — АДАПТОГЕНЫ В МЕХАНИЗМАХ РЕГУЛЯЦИИ ЖИЗНЕДЕЯТЕЛЬНОСТИ КЛЕТКИ
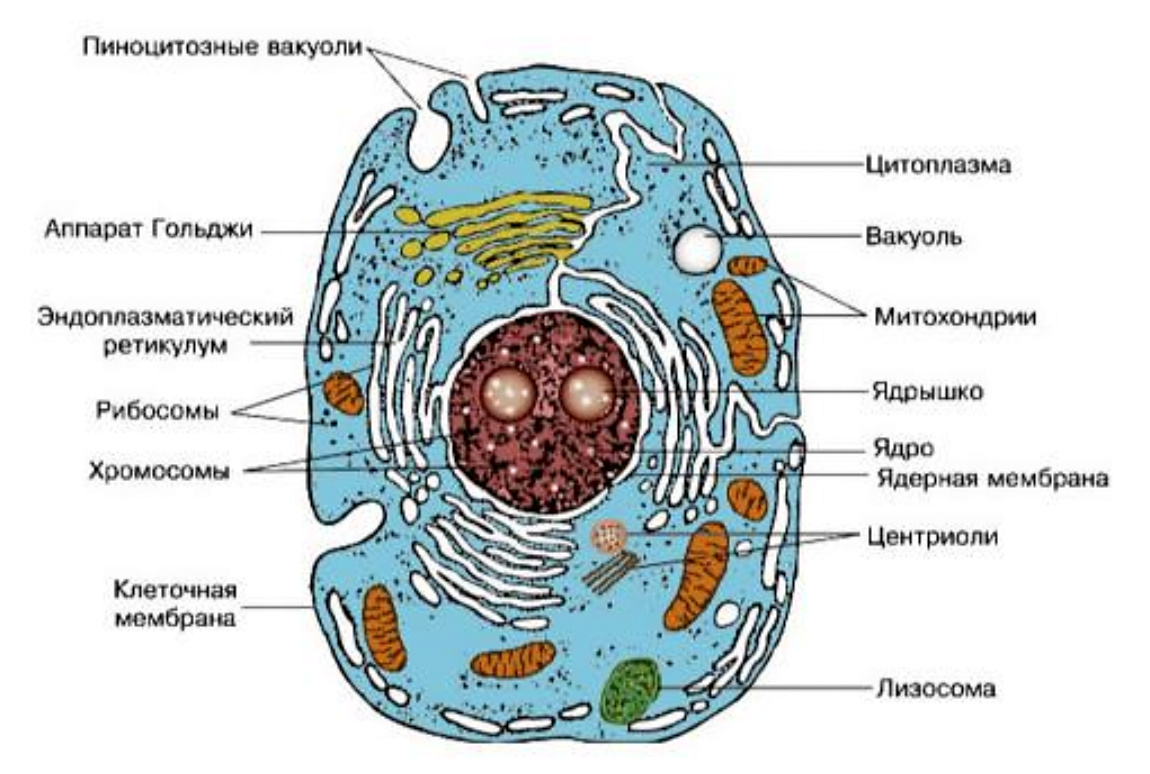
Начать обобщение наших знаний об адаптивных механизмах живой материи необходимо с элементарной ячейки живой материи-клетки. На приведенном рисунке можно видеть ряд важнейших структур клетки. Важнейшими органеллами, во многом определяющими адаптивные перестройки клетки, являются митохондрии, в которых протекает окисление поступающих в клетку питательных веществ (углеводы и жиры). В результате чего образуется энергия АТФ, выделяется углекислый газ и вода. Окислительно-восстановительные процессы (ОВП) в митохондриях протекают при интенсивном использовании кислорода. (Рис.2)
На сегодня установлено, что для процессов жизнедеятельности организмов, а, соответственно, и для защитно-адаптивных механизмов необходима энергия, которая накапливается в клетках в виде макроэргического соединения АТФ (аденозинтрифосфат), или в виде ГТФ (гуанизинтрифосфат). В дальнейшем АТФ используется для самых разнообразных структурно-функциональных перестроек и динамической работы и других функциональных процессов клеток и тканей. ГТФ используется в процессах биосинтеза, то есть в построении новых биологических структур [235,156,36,34].
АТФ можно рассматривать как первый медиатор-гормон, появившийся на заре зарождения живых организмов [352]. Имея в своем составе адениловый нуклеотид и остатки фосфорной кислоты АТФ обладает многочисленными регуляторно-защитными функциями начиная от способности аккумулировать и отдавать энергию и выполняет различные регуляторные функции как на уровне мембран, так и на уровне цитозоля клетки. Соотношение АТФ-АДФ-АМФ влияет на многие стороны жизнедеятельности клетки, прежде всего ее энергетические функции. Важное регуляторное значение имеют производные АТФ и ГТФ циклические нуклеотиды — ЦАМФ и ЦГМФ, регулирующие пластические и функциональные процессы в клетке. Для образования энергии в клетках и тканях необходимо поступление в них достаточного количества кислорода. Следует отметить, что большинство организмов используют в своих окислительно-восстановительных процессах в качестве окислителя кислород (аэробные организмы). Но есть организмы, живущие в бескислородной среде и использующие в своих окислительно-восстановительных процессах другие окислители, например серу, азот и некоторые кислоты. Но наиболее эффективное образование энергии и возможность активного функционирования отмечается именно у аэробных организмов, использующих кислород. То есть в филогенетическом плане это более совершенные и адаптивные организмы [34, 36, 170, 235, 291, 358, 359,378,473,474].
Рис. 2. Схема строения живой клетки:
Процессы образования энергии являются неспецифическими, т.е. необходимы для всех организмов в их адаптивных перестройках и ничем у аэробных организмов не отличаются. Они протекают абсолютно одинаково у человека, животных, простейших и бактерий, использующих кислород.
Окислительно-восстановительные процессы в клетках происходят в нескольких формах:
1. Окисление поступающих с пищей углеводов, жиров и белков (аминокислот, которые могут превращаться в углеводы в процессе глюконеогенеза) происходит в митохондриях клеток при участии группы ферментов, объединенных в так называемый цикл трикарбоновых кислот или цикл Кребса. В регуляции функции митохондрий ключевое значение имеют системы, сопрягающие азотистый и энергетический обмен. Процесс переаминирования и дезаминирования аминокислот и включения их в энергетические процессы или напротив, образования аминокислот, происходят с участием трансаминаз аланина и аспарагиновой кислоты, а так же фермента — глутаматдегидрогеназы. Кроме того, имеются шунтовые механизмы с участием аспарагиновой кислоты и аланина, сопрягающие процессы метаболизма в митохондриях и цитозоле клеток, т.е. окислительно-восстановительные процессы тесно связаны с обменом аминокислот и находится под контролем соотношения аминокислот — адаптогенов. В условиях повышенных нагрузок на организм важное энергетическое значение имеют аминокислоты, образовавшиеся вследствие усиления протеолитических процессов из которых, преимущественно в печени, образуется глюкоза — процессы глюконеогенеза. Строго говоря, вторая фаза стресс реакции на метаболическом уровне во многом сводится к усилению протеолитических процессов за счет выброса глюкокортикоидов и обеспечения организма необходимой энергией за счет реакций глюконеогенеза [145,86].
Важнейшим веществом, которое образуется при таком окислении в митохондриях является НАДН2 (никотинамиддинуклеотид восстановленный). В такой промежуточной форме накапливается энергия окисления перечисленных выше питательных веществ. Затем, от НАДН2 отщепляется водород и электроны, с помощью дыхательной цепи (группы окислительно-восстановительных ферментов, находящихся в митохондриях), то есть там же, где протекает цикл Кребса. В результате окислительно-восстановительных процессов в дыхательной цепи образуется универсальное энергетическое соединение АТФ (аденозинтрифосфат), обеспечивающее большинство энергоемких процессов в тканях организма, углекислый газ и вода, которые выделяются из организма легкими и почками.
Необходимо отметить, что НАД, так же как и АТФ, является важнейшим защитно-регуляторным метаболитом, обеспечивающим регуляцию как энергетических, так и пластических процессов за счет способности к окислению и восстановлению. Соотношение восстановленных и окисленных его форм во многом определяет механизмы регуляции. Присоединение фосфата дает другой важный защитно-регуляторный метаболит — НАДФ. Таким путем спектр защитно-регуляторных возможностей этих метаболитов существенно возрастает. Необходимо отметить, что как пуриновые, так и пиримидиновые нуклеотиды, входящие в состав АТФ и НАДФ являются производными нескольких аминокислот: аспарагиновой, глицина, глутамина и наличие этих аминокислот в клетках во многом влияет на образование перечисленных нуклеотидов и их отношение [352, 34, 36, 170, 235, 291].
2. Другой группой окислительно-восстановительных процессов в клетке является так называемый пентозный цикл, протекающий в цитоплазме клетки. В этом цикле окисляются углеводы (глюкоза) с образованием НАДФН2 (никотинамиддинуклеотидфосфат восстановленный). Этот процесс не требует кислорода, а образующийся НАДФН2 необходим для поддержания процессов биосинтеза вновь образующихся структур клетки, особенно при их делении и росте. Также большое значение пентозный цикл имеет в обеспечении обезвреживания (детоксикации) попадающих в организм чужеродных веществ (ксенобиотиков) или структур тканей и клеток, подвергающихся повреждению, которые необходимо выводить из организма (эубиотики). Ведущее значение в детоксикации попадающих в организм чужеродных веществ (ксенобиотиков) или подлежащих выведению продуктов межуточного обмена (эубиотиков) имеет печень, хотя в определенной степени такие же процессы протекают, например, в легких, коже, клетках крови и т.д.
В регуляции многих сторон жизнедеятельности клетки большое значение имеют регуляторные метаболиты, имеющие в своем составе SH группы, прежде всего это глутатион. Поддержание этих групп в восстановленном состоянии обеспечивается системами образования НАДФН2 (пентозный цикл, работа маликфермента) и образование аскорбиновой кислоты в соединительнотканных структурах организма. Функции этих систем тесно взаимосвязаны и, как показано в наших исследованиях, регулируются метаболитами обмена глутаминовой кислоты: оксипролин, аргинин и ряд других, т.е. так же зависимы от обмена аминокислот.
3. Еще одной группой окислительно-восстановительных процессов являются процессы обезвреживания ксено- и эубиотиков ферментами, располагающимися на гладком эндоплазматическом ретикулуме, то есть системе оксигеназ смешанных функций (ОСФ) находящихся на мембранах, как бы сложенных в цитоплазме клетки, то есть окислительно-восстановительных процессов с использованием кислорода протекают и в цитоплазме клетки. Это необходимо для инициации детоксикационных процессов при окислении чужеродных веществ (ксенобиотиков), попадающих в клетку, или обезвреживании токсичных продуктов, образующихся в тканях в ходе обмена веществ (эубиотики). В дальнейшем, после окисления, эти вещества соединяются с обезвреживающими метаболитами, такими, как глутатион, некоторые аминокислоты, и ряд других веществ с дальнейшим выведением из клеток, тканей и организма.
Имеются еще некоторые группы окислительно-восстановительных процессов, например, прямое окисление липидов в пероксисомах.
4. Наконец в цитоплазме клеток протекает наиболее древний процесс образования энергии – гликолиз, который не требует кислорода и расщепляет исключительно глюкозу. В гликолизе энергии АТФ образуетсмя в 18 раз меньше, чем при аэробном окислении и может накапливаться молочная кислота при недостаточно эффективных процессах аэробного окисления и недостатке кислорода.
Для нормального протекания окислительно-восстановительных процессов необходимы следующие условия: как уже отмечалось, обеспечение в достаточном количестве тканей организма кислородом с одной стороны, с другой необходима адекватная данной нагрузке антиоксидантная защита. В случае, если эти процессы сбалансированы, то адаптивные системы организма способны выводить его на уровень, устойчивый к тому или иному повреждающему воздействию. Если необходимого соответствия в этих процессах нет, то возникает либо гипоксическое состояние (нехватка кислорода), либо состояние пониженной антиоксидантной защиты. В том и в другом случае снижается образование носителей энергии в клетке АТФ и ГТФ, нарушается ее функция, а затем функции тканей и системы в целом. Длительное воздействие повреждающего фактора с понижением антиоксидантной защиты приводит к развитию «окислительного стресса» с повреждением мембран и других структур клеток и тканей, с нарушением их функций, а в случае продолжения повреждающего воздействия, к их гибели [19, 20, 179, 180, 65, 29, 30, 179, 181, 412, 457, 487, 555, 558, 522, 530, 589].
Кислород — это мощный окислитель, который является достаточно агрессивным веществом, способным повреждать мембраны и другие структуры клеток, контактирующих с ним. Еще более агрессивными являются промежуточные продукты, образующиеся из кислорода в ходе ОВП. К таким продуктам относят супероксидный анион, синглетный кислород, перекись водорода и ряд других веществ, способных атаковать окружающие мембраны и структуры клеток. В норме процессам избыточного образования активных форм кислорода (АФК), токсичных для клетки, препятствует антиоксидантная защита (АОЗ) клеток и тканей. Эта система включает в себя несколько белков-ферментов (супероксиддисмутаза, каталаза, пероксидаза, глютатионредуктаза и глютатионпероксидаза), а также различные вещества, способные препятствовать повреждающему воздействию кислорода и его активных форм. К таким веществам относят витамины А, Е, К и С, многие гормоны и биологически активные вещества. А также ряд других простых и сложных соединений. Однако необходимо отметить, что многие из перечисленных метаболитов и сложных соединений при интенсивном и часто повторяющемся стрессовом воздействии на ткани и клетки утрачивают антиоксидантные возможности, а зачастую, и сами становятся прооксидантами и агрессивными веществами, способными повреждать мембранные структуры. Например, гормоны (катехоламины), биологически активные амины (серотонин, гистамин), витамины (витамин С) и ряд других веществ. И только аминокислоты, особенно серосодержащие (цистеин, таурин), а также содержащие повышенное количество азота (глутамин, аргинин, лизин) до конца выполняют свою антиоксидантную функцию. Еще более мощный антиоксидантный защитный эффект оказывают некоторые производные перечисленных аминокислот: глутатион, полиамины, а также ряд других веществ, содержащих в своем составе несколько отмеченных выше аминокислот (олигопептиды) [197, 198, 199, 200, 201, 202, 22, 23, 25, 117, 126, 185, 223, 304, 334, 393, 469, 481, 587].
Другой важнейшей структурой клетки — является ядро, в значительной мере определяющее специфический, присущий только этому организму ответ на повреждающее воздействие. В ядре сосредоточена генетическая информация в виде спирали ДНК, находящаяся в хромосомах. Эта информация может воспроизводиться за счет информационных РНК, которые передают заложенную в генах информацию на рибосомы клетки (расположенных на мембранах эндоплазматического ретикулума — шероховатый эндоплазматический ретикулум). На рибосомах происходит биосинтез белков- ферментов, необходимых для протекания любых обменных процессов в организме. Рибосомы — состоят из рибосомальной РНК и белков, они являются своеобразными фабриками по биосинтезу вновь образующихся белков. В этом процессе используются 20 основных аминокислот, которые как кирпичики складываются в первичную структуру белка. Затем силы электрофизического и химического взаимодействия скручивают эту последовательность в спираль — вторичную структуру белка, которая затем складывается под воздействием этих же сил, а также физико-химических факторов окружающей среды в третичную структуру — собственно сам белок. Далее такие субъединицы могут соединяться в функционально-активные белковые объединения, например, некоторые ферменты, состоящие из нескольких субъединиц. В последовательности аминокислот, а также во всех последующих структурах белковой молекулы заложена колоссальная информация, которая реализуется в ходе различных обменно-адаптивных превращений [34, 235, 36, 43, 155, 175].
Считается, что в регуляции воспроизводства наследственной хромосомной информации большое значение имеют гормоны (соматотропный, глюкокортикоиды, инсулин). Кроме того, имеется обширная, но противоречивая литература о влиянии на генетический аппарат количества SH групп, активных форм кислорода, свободных радикалов. Однако, какой-либо определенной и убедительно обоснованной концепции о регуляции генетического аппарата продуктами метаболизма до сих пор не существует [117, 291]. Большинство работ, посвященных функции генетического аппарата и биосинтеза белков в клетках выполнены на Е.соli. У многоклеточных организмов эти процессы идут значительно сложнее, и многие стороны их еще мало изучены. Поэтому возможности эффективно воздействовать на эти процессы пока нет [352, 142].
Исходя из данных литературы [22, 193, 194, 195, 196, 197, 198, 199, 200, 201, 202] и результатов наших исследований, аминокислоты-адаптогены, влияющие на биосинтез таких важных регуляторов метаболизма как пуриновые и пиримидиновые нуклеотиды, глутатион и другие биологически активные олиго- и полипептиды, например, тафтсин, холецистокинин, гистоны, полиамины, находясь в определенном соотношении, соответственно, влияют на функцию генетического аппарата клетки. Иными словами, они могут сопрягать неспецифическую и специфическую фазы стресс-реакции с последующим образованием необходимых белков — ферментов, выводящих организм из фазы тревоги в фазу резистентности.
Так, белки выполняют регуляторную, рецепторную, защитно-опорную, а также другие функции, обеспечивающие специфический ответ клетки на внешние воздействия и изменение обменных процессов под этими воздействиями. Наибольшее значение для выполнения структурно-опорной функции имеют белки соединительной, костной ткани, сосудов, нервных волокон. Специфические функционально-морфологические особенности организма во многом определяются иммунной системой, в которой образуются белки — антитела и антигены. Иммунная система выполняет защитную функцию по отношению к чужеродным элементам (например, возбудитель инфекционных заболеваний) устраняя их из организма, а также элиминируя собственные измененные и поврежденные клетки и ткани.
Белки играют значительную роль в регуляции обменных процессов организма, так они встраиваются в мембраны клеток, определяя их функциональное значение. Работа митохондрий, ядра, рибосом, обезвреживающих механизмов на гладком эндоплазматическом ретикулуме выполняются соответствующими белковыми структурами.
Как ферменты, белки обеспечивают протекание всех обменных процессов в организме. В последовательности аминокислот и их сборке во вторичную, третичную, четвертичную структуру, заложена колоссальная информация, которая накапливалась миллиарды лет в процессах эволюции живой материи. Достаточно сказать о том, что протекание реакций обмена веществ без наличия белков-ферментов потребовало бы температуры в сотни и тысячи градусов, высокого давления и других, несовместимых с физиологическим уровнем клетки условий. Поэтому живой материи на планете. Однако, помимо того, что аминокислоты выстраиваются в определенную последовательность в белках, или более коротких молекулах — полипептидах или олигопептидах, они также находятся в свободном состоянии во всех клетках и тканях организма и выполняют важнейшую регуляторную функцию. Как мы уже отмечали, соотношение этих свободных аминокислот строго постоянное и меняется лишь под воздействием интенсивных повреждающих факторов или значительных адаптивных перестройках организма [158, 126, 235, 143]. Все это позволяет заключить, что соотношение аминокислот как в молекулах белков или полипептидов, так и в свободном состоянии является (в терминологии пифагорейцев) сакральной геометрией живой материи или тем математическим языком, который регулирует эту материю, через который окружающая среда взаимодействует с организмами или популяциями этих организмов. Таким образом, аминокислоты, создавая структуры белка (первичную, вторичную, третичную, четвертичную), а также создавая различные соотношения в клетках и тканях в живой материи, во многом определяют ее энергетически-адаптивный потенциал и, изменяясь в результате внешних воздействий, эти соотношения регулируют защитно-адаптивные механизмы.
1.3 ВАЖНЕЙШИЕ ЗАЩИТНО-АДАПТИВНЫЕ СИСТЕМЫ ОРГАНИЗМА
Для контроля за адекватным протеканием адаптивных перестроек к изменениям окружающей среды в организме имеются неспецифические защитно-адаптивные системы.
1. Нейроэндокринная. Эта система включает в себя мозг, в котором выделяют кору и подкорковые структуры, а также продолговатый и спинной мозг и железы внутренней секреции (гипофиз, щитовидная железа, надпочечники, поджелудочная железа, половые железы).
В нейронах в условиях стресс-воздействия образуется большое количество РНК, биологически активных белков, олиго и полипептидов (в настоящее время известно более 600 биологически активных пептидов мозга способных регулировать функцию различных адаптивных систем организма), что во многом определяет формирование процессов памяти [22, 198]. Значительное усиление метаболических процессов сопряжено с избыточной энергообеспеченностью, которая необходима для реализации адаптивных процессов [22, 197, 198, 350, 352, 268]. При этом в головном мозге в больших количествах образуются такие метаболиты как аммиак, мочевина, глутаминовая и аспарагиновая аминокислоты. Все эти вещества являются мощными регуляторами защитно-адаптивных механизмов [158, 197, 22].
Важнейшее значение в регуляции нейроэндокринной системы, особенно ее высших отделов (кора головного мозга и подкорковые структуры), имеют некоторые аминокислоты-адаптогены. К возбуждающим аминокислотам — медиаторам относят глутаминовую и аспаргиновую кислоты и ряд их производных, а также триптофан и фенилаланин, терозин и их производные (катехоламины (КХА), гормоны щитовидной железы — тироксин и другие). К тормозным аминокислотам — медиаторам относят гамма-аминомасляную кислоту — ГАМК, таурин, глицин [22, 263, 68, 98, 595, 88, 89, 150, 264, 360, 372, 373, 441, 443, 407, 410, 420, 428, 431, 454, 456, 468, 469, 486, 485, 497, 503, 541, 551, 549, 552, 528, 572, 567, 578, 579, 590, 592, 596, 597]
Нейроэндокринная система имеет главенствующее значение в адаптивных перестройках организма, как к воздействиям внешней среды, так и внутренним изменениям организма. На основе сигналов об изменении ситуации во внешней или внутренней среде формируется программа действий, реализуемая посредством как опорно-двигательного аппарата, так и посредством изменений со стороны внутренних органов. То есть информация передается на подкорковые структуры (гипоталамус, гипофиз). В гипоталамусе вырабатываются биологически активные вещества — рилизинг-факторы, которые поступают в гипофиз. Далее в гипофизе вырабатываются тропные гормоны, например адренокортикотропный гормон, который имеет важнейшее значение в реализации механизмов стресса. Он стимулирует выработку надпочечниками важнейших гормонов стресса: катехоламинов (КХА) и глюкокортикоидов (ГК). Катехоламины значительно усиливают расщепление углеводов из гликогена («депо» углеводов в клетках), а также мобилизуют жиры из их «депо» в органах и подкожно-жировой клетчатке. В результате в крови резко повышается уровень углеводов (глюкоза) и жиров (жирные кислоты), которые поступают в клетки и интенсивно окисляются в их энергетических центрах-митохондриях. Здесь образуется энергия в виде АТФ, необходимой для стимуляции адаптивно-защитных механизмов и повышения устойчивости к повреждающему стресс-воздействию [86, 22, 47, 63, 64, 62, 91, 96, 97, 106, 213, 246, 279, 332, 398, 416].
Глюкокортикоиды способствуют расщеплению белков в ряде тканей организма, подвергающихся разрушающему воздействию стрессового фактора: лимфоидная ткань, соединительная, мышечная, жировая ткань, некоторые ткани желудочно-кишечного тракта, и некоторые другие. Смысл такого разрушения состоит в резком увеличении аминокислот-адаптогенов для стимуляции защитных механизмов, противостоящих процессам повреждения и стимулирующих восстановительные биосинтетические процессы (реконструктивный протеолиз), что в целом повышает устойчивость организма к повреждающему фактору (Покровский А.А. «Лизосомы» 1976). Но имеются данные, что различные организмы различно и реагируют на воздействие глюкокортикоидов. Могут быть глюкокортикоид+ или – организмы [84]. В наших исследованиях установлено, что реконструктивный протеолиз более выпажен у глюкокортикоид «-» морских свинок, чем у глюкокортикоид «+» крыс, что отмечено в схеме №1
2. Сердечно-сосудистая система. Эта система включает в себя сердце, выполняющее механическую функцию и сосуды различного диаметра. Эта система выполняет функцию транспорта кислорода (в эритроцитах), от легких к тканям, где поступающий кислород включается в ОВП. А также транспорта других необходимых для обеспечения жизнедеятельности веществ (углеводов, жиров, аминокислот, витаминов, электролитов и других).
3. Дыхательная система: верхние дыхательные пути (бронхи) и легкие, которые обеспечивают проникновение кислорода из вдыхаемого воздуха в кровеносные сосуды, а затем в эритроциты.
Имеются сведения, что в регуляции этих систем на метаболическом уровне большое значение имеет образование оксида азота и соответствующие ему ферменты. Образование оксида азота и его регуляторное значение на защитно-адаптивные системы организма тесно связанные с обменом аммиака, глутаминовой кислоты, аргинина и мочевины. В тканях имеются различные NO синтетазы (Е-эндотелиальные, Н-нейрональные, I-индуцибельная), которые определяют наработку оксида азота в эндотелии, нейронах и фагоцитирующих клетках. С определенным уровнем оксида азота связаны функциональные возможности этих систем [257, 123, 130, 140, 449, 417, 470, 535].
4. Желудочно-кишечный тракт обеспечивает поступление пищевых веществ и их расщепление до метаболитов, способных всасываться через кишечную стенку в кровь и транспортироваться в виде углеводов, жиров, аминокислот к тканям в качестве необходимого топлива для протекания окислительно-восстановительных процессов и процессов биосинтеза.
Схема 1. 1. Метаболические особенности морской свинки и крысы при воздействии на них МБТ и мутагенных ксенобиотиков типа ПАУ:
В желудке и кишечнике вырабатываются биологически активные пептиды такие как холецистокинин, серотонин, гистамин и ряд других оказывающих влияние на эмоционально волевую сферу, процессы памяти, сна. Наработка этих веществ влияет на процессы интеграции между адаптивными система организма. Печень является своеобразной биохимической лабораторией организма, образующей большое количество биологически активных адаптогенов.
5. Кроветворная — сосредоточена в трубчатых и плоских костях в виде костного мозга и необходима для постоянного образования клеток крови: лейкоцитов, эритроцитов, тромбоцитов, выполняющих различные защитно-адаптивные функции в организме.
6. Иммунная система включает в себя железы регуляции иммунитета (вилочковая железа — тимус), а также периферические ткани иммунной системы (селезенка, лимфатические узлы различных органов, тканей и сосудов).
Имеются данные, что в регуляции этих систем так же имеют большое значение аминокислоты, принимающие участие в реализации регуляторных механизмов (цистеин, таурин, глутаминовая и аспарагиновая аминокислоты, метионин, аргинин, пролин) [158, 32, 22, 239, 10, 26, 82, 88, 124, 117, 126, 143, 68, 224, 226, 393, 417, 431, 443, 465, 476, 493, 504, 510, 511, 526, 533, 534, 540, 541, 552, 562, 566, 567, 576, 578, 593, 595, 598].
7. Антиоксидантная система — система, отвечающая за предотвращение повреждающего воздействия кислорода, его промежуточных метаболитов на важнейшие структуры клетки, прежде всего мембраны (на чем мы останавливались выше).
8. Выделительная система это система обезвреживания вредных веществ поступающих в организм извне (ксенобиотики), а также поврежденных и измененных структур и молекул собственно организма — эубиотиков. Начинается такое обезвреживание в метаболических системах клетки.
Первая и ключевая реакция — обезвреживания в метаболических системах аммиака, образующегося в высокофункциональных тканях, прежде всего в нейронах головного мозга. Обезвреживание происходит при связывании аммикака глутаминовой кислотой с образованием глутамина и выведением его за пределы клетки.
На мембранах гладкого эндоплазматического ретикулума (ГПР) ксенобиотики или образующиеся в организме вещества подлежащие обезвреживанию, подвергаются окислению в системе оксигеназ смешанных функций (ОСФ), а затем связыванию полученных метаболитов с некоторыми аминокислотами (таурином, глицином, глютамином) а также некоторыми специально синтезируемыми веществами (глутатионом, уридиндифосфоглюкуроновой кислотой (УДФГК) и фосфоаденозинфосфосульфатом (ФАФС) и выводятся из клетки через аппарат Гольджи, представляющий из себя конечные отделы эндоплазматического ретикулума. Эти процессы наиболее интенсивно протекают в клетках печени, легких и менее интенсивно в коже, клетках крови и клетках других тканей. Затем обезвреживаемые вещества в виде соединений с перечисленными выше метаболитами выделяются в кровь и далее с мочой, потом и другими выделениями выделяются из организма [85, 168, 223, 225, 227].
Отмеченные неспецифические защитно-адаптивные механизмы, рассматриваемые нами на уровне отдельных организмов универсальны, но живая материя входит в единую систему-биоценоз, который также можно рассматривать, как своеобразный организм с присущими ему защитно-адаптивными системами и механизмами развития. Эти особенности реализуются на видовом, межвидовом, популяционном и других уровнях биоценоза. Существует различные биологические цепи, определяющие как качество жизни, так и самое существование того или иного вида. Так, в пищевой цепи, травоядные зависят от наличия растительной пищи, хищники зависят от количества травоядных. Погибающие организмы как травоядных, так и хищников служат пищей для простейших и бактерий, преобразующих их останки в азотистые удобрения, и увеличивают массу растительной пищи и т.д. Другим примером биологической цепи может быть трансгенный перенос между различными видами животных и человеческих популяций, осуществляемый микроорганизмами (простейшие, бактерии, вирусы). Микромир имеет очень высокий потенциал адаптивных перестроек и очень чутко реагирует на изменение окружающей среды (как глобальные космические воздействия, так и отрицательные последствия деятельности человека), поэтому трансгенный перенос осуществляется микроорганизмами, переносящими метаболически-генетическую информацию как между отдельными особями, так и между различными видами, что является важнейшим адаптивным механизмом биоценоза [378, 507].
Существуют разнообразные формы симбиотических (взаимовыгодных) отношений между организмами. Так, некоторые виды птиц уничтожают паразитов на теле и в ротовой полости китов, крокодилов, бегемотов. Последние, в свою очередь, служат защитой для этих птиц.
Еще большее значение для регуляции защитно-адаптивных систем макроорганизмов имеет микрофлора: бактерии, простейшие, спирохеты, грибы, вирусы и другие ее представители. Микроорганизмы распространены повсеместно, во всех слоях биосферы и контактируют со всеми населяющими ее организмами, активно воздействуя на их защитно-адаптивные системы. Так хорошо известен факт, что у человека на поверхностях, контактирующих с внешней средой, находится большее количество микроорганизмов, чем его собственных клеток, составляющих различные органы. И все эти микроорганизмы выполняют какую-то свою симбиотическую функцию. Например, микрофлора кишечника, вырабатывает основное количество необходимых для организма витаминов группы В — В1, В6, В12 и ряд других важных компонентов для нормального протекания обменных процессов [378,43,235].
В случае гибели нормальной микрофлоры, например при некоторых заболеваниях желудочно-кишечного тракта, развивается дисбактериоз с чрезмерным размножением патогенных бактерий, грибов. Нарушается обеспечение организма витаминами, в организм попадают токсичные вещества, может развиться тяжелое нарушение обмена веществ, вплоть до гибели организма, что чаще отмечается у детей. Введение необходимых микроорганизмов (бифидумбактерии, лактобактерии, кишечная палочка) препятствуют развитию и прогрессированию заболевания.
В наших исследованиях [236, 237, 238, 239] было установлено, что микобактерии туберкулеза существенно снижают мутагенное воздействие ароматических ксенобиотиков на экспериментальных животных.
Согласно симбиотической концепции [378], важнейшие энергетические структуры клетки-митохондрии являются потомками бактерий, которые взаимодействовали с первичными многоклеточными оранизмами в анаэробный период биосферы. Это повышало адаптивные возможности тех и других. В дальнейшем, произошло их слияние в единую систему. Имеются точки зрения, что помимо митохондрий тканей, микротрубочки нейронов мозга так же имеют бактериальное происхождение. Кроме того, метаболический аппарат оксигеназ смешанных функций в наибольшей степени выраженный в печени, так же, возможно, имеет бактериальное происхождение. Однако, если бактерии, послужившие родоначальниками митохондрий современных организмов, обладали преимущественно ферментными системами, способными к метаболическим превращениям с участием кислорода, то бактерии или грибобактерии, давшие начало оксигеназам смешанных функций, имели мощные ферментные системы, включавшие в себя цитохром Р 450. Последние в большей степени были способны интенсивно метаболизировать ксенобиотики типа ПАУ (полициклических ароматических углеводорода).
Определенным подтверждением такой концепции можно считать отсутствие цитохрома Р-450 и системы ОСФ почти на всех ступенях эмбрионального развития большинства животных и человека. Необходимо отметить, что каждый индивидуум проходит все ступени филогенеза в своем онтогенезе и только после рождения и по мере взросления организма количество цитохрома Р-450 резко увеличивается в тканях (печени, легких, кожи) [85, 227, 168].
Примечательно, что образование стероидных гормонов в надпочечниках и половых железах, для образования которых также необходим цитохром Р-450, также начинается после рождения плода, т.е. после резкого усиления аэробных процессов в организме [86]. Все это наводит на мысль о том, что образование стероидных гормонов — своеобразный эндогенный механизм регуляции, который в какой-то мере дополняет или конкурирует с регуляторными воздействиями экзогенных биорегуляторов – полициклических ароматических углеводородов (ПАУ). Регуляция эта может быть связана с тем, что стероиды, так же как и ПАУ, которыми они по сути дела и являются, так как в их основе лежит молекула циклопентанпергидрофенатрена, обезвреживаются или подвергаюся метаболическим превращениям по тем же путям, что и ПАУ. Или иными словами ПАУ можно рассматривать как экзогормоны, а образующиеся в организме метаболиты сходные с ними по строению как эндогормоны или ксенобиотики и эубиотики.
Эти вещества легко проникают в липиды организма, изменяя их свойства, т.е. свойства мембран [85, 168, 227]. Подвергаясь обезвреживанию в системе оксигеназ смешанных функций (ОСФ), эти вещества могут давать метаболиты, ковалентно связывающиеся с нуклеиновыми кислотами и другими нуклеофильными и активными центрами биологических молекул. Это обуславливает мутагеные и канцерогенные воздействия ПАУ т.е., вызывающие мутации — изменение генетической информации и развитие патологических процессов вплоть до опухоли, нарушение окислительно-восстановительных процессов клеток и ряд других нарушений метаболизма, связанных с образованием активных форм кислорода и агрессивных метаболитов ПАУ и перекисного окисления липидов (ПОЛ).
Согласно представлениям, сформулированным учеными в последние годы [501, 507] микромир играет исключительную роль в интеграции биоценоза. Установлено, что микроорганизмы могут трансформироваться друг в друга, утрачивая свою прежнюю специфику и приобретая новые свойства. Ежедневно микромир обменивает до 15% своей генетической информации. При этом он осуществляет трансгенный перенос этой информации, проникая в различные макроорганизмы и взаимодействуя с их генетическими структурами. И мы можем добавить, что микромир взаимодействует и с метаболическими защитно-адаптивными системами макроорганизмов, таким путем интегрируя их на уровне видов, популяций, биоценоза в единые защитно-адаптивные системы.
Согласно концепции Чижевского А.Л. («Эпидемические катастрофы и периодическая деятельность солнца» М. 1930 ), микромир имеет большое значение для развития видов животных и популяций человеческого общества и соответственно цивилизации. Так эпидемии и пандемии таких опасных заболеваний как чума, оспа, холера, туберкулез имеют определенную цикличность во времени и влияют на процессы цивилизационного развития.
1.4 РОЛЬ АМИНОКИСЛОТ — АДАПТОГЕНОВ В МЕХАНИЗМАХ РЕАЛИЗАЦИИ СТРЕССОВОЙ РЕАКЦИИ
В 30-40 годах прошлого столетия Г.Селье была разработана теория стресса[293]. Она описывает универсальное свойство живой материи при воздействии на нее различных повреждающих факторов. Но несмотря на большие успехи в области биохимии, физиологии, молекулярной генетики и других направлениях, достигнутых с того времени, что якобы, позволяет даже клонировать не только животных, но и человека и разрабатывать другие методы воздействия на генотип, во многом эти попытки сродни средневековой алхимии.
Однако если алхимик подвергал риску только собственную голову, в данном случае опасность подстерегает человечество в целом, так как отдаленные результаты этих экспериментов практически предсказать нельзя [142]. В механизмах адаптации и регуляции адаптивных систем остается много неясного.
Во многом остается непонятно, как осуществляется координация между важнейшими гомеостатическими системами организма и генетическим аппаратом его клеток, не известны медиаторы, которые модулируют функцию генетического аппарата. Во всяком случае, в наших руках пока нет средств, позволявших бы активно вмешиваться в механизмы стрессового воздействия без отрицательных последствий для организма [357].
Несмотря на то, что теория стресса подробно разработана и имеет многочисленных сторонников, по мере углубления знаний о механизмах повреждающего воздействия различных факторов на организм и о различных типах реагирования на эти воздействия, появляется все больше противоречий в трактовке механизмов адаптивного ответа на стресс воздействие. Это может быть связано как филогенетически обусловленными межвидовыми отличиями, так и вызванными различными патологическими состояниями нарушениями адаптивных механизмов. Более того, внутри каждого вида найдены существенные отличия в механизмах реагировании.
Такие известные физиологи, занимавшиеся проблемой адаптации как Л.А.Орбели, П.Д.Горизонтов, В.П.Казначеев, Ф.З.Меерзон, Л.Е.Панин и многне другие, не отрицая понятия стресс, предлагают свои схемы ответа организма на повреждающее воздействие. В результате возникает пестрая картина, которая не позволяет выработать единую теорию и алгоритм подходов для объяснения понятий стресс и адаптация и их взаимодействия.
Однако иногда спор доходит до отрицания самого понятия стресса, имеется обширная литература, в которой расматривается различные механизмы стрессового воздействия [3, 5, 14, 15, 31, 46, 47, 64, 67, 80, 81, 91, 97, 105, 106, 118, 119, 120, 133, 150, 153, 159, 176, 191, 192, 222, 268, 279, 321, 322, 346, 350, 363, 371, 375, 381, 384, 385, 386, 394, 476, 522, 530, 535, 592]
Другой важной проблемой является наличие специфических и неспецифических реакций при стрессе и их взаимоотношения. Еще сам Г.Селье подчеркивал, что основные проявления стресс реакций не специфические, однако некоторые авторы подвергают сомнению неспецифический характер, так как каждый стимул имеет свой характерный ответ. Все это существенно затрудняет как понимание, так и коррекцию последствий перенесенных воздействий.
В работах отечественных исследователей [240, 199, 244] обоснованно выделяются как неспецифические, так и специфические факторы стресса.
Причем первые — удел фазы тревоги, когда включаются реакции нейроэндокринной системы, мобилизация жиров, углеводов, аминокислот, катаболические процессы, что сопровождается образованием эндогенных стимуляторов, резко повышающих процессы энергетики и запускающих каскад метаболических процессов, направленных на повышение резистентности организма и выход его на адаптивный уровень. В исследованиях Ф.З. Меерзона показано, что неспецифические процессы в определенной степени формируют специфичный ответ, который во многом заключается в формировании памяти на уровне генетического аппарата клеток. Наибольшей емкостью этой памяти обладают клетки нейроэндокринной системы, прежде всего нейронов. В которых резко возрастает количество функционально биологически активных ДНК, РНК, олиго и полипептидов и аминокислот — нейромедиаторов. Высказывается мысль о том, что образующиеся в тканях мозга метаболиты — адаптогены непосредственно могут включаться в метаболические процессы на уровне поврежденных тканей (глутаминовая кислота, ГОМК, полипептиды и др. [22,198,352]. Таким путем нервная система может оказывать регуляторное воздействие не только путем генерации нервного импульса, но и непосредственно за счет выхода метаболитов в кровеносное русло. В тоже время надо признать, что факторы, связывающие неспецифическое и специфическое звено стресса не найдены, что является слабым звеном теории стресса, и нет достоверных, научно обоснованных подходов для воздействия на реализацию стресс реакции в нужном нам направлении.
Мы же полагаем, что таким фактором является характер деятельности, в той или иной степени определяющий деятельность. Здесь надо обратить внимание на то, что по нашим данным характер деятельности организма у спортсменов представляющих разные виды спорта имеется отчетливая разница в характере метаболизма, выраженная в различии количества и качества аминокислотного состава плазмы крови. Этот факт указывает на преобладание разных фаз стресса у представителей разных видов спорта.
Биологический смысл первой фазы стресса состоит в «смывании» текущей доминанты в коре головного мозга и усиление энергообеспечения организма с целью формирования новой доминанты, соответствующей новым условиям, возникновение которых и вызвало стресс-реакцию. Биологический смысл второй фазы стресса состоит в том, что он оставляет структурный след адаптации. Происходят изменения на генетическом уровне, усиливается биосинтез белков — ферментов, ответственных за перестройку метаболических систем, лежащих в основе функционирования важнейших адаптивных систем: нейроэндокринной, дыхательной, сердечно-сосудистой, выделительной, детоксикационной, антиоксидантной.
Согласно полученных Паниным Л.Е. и соавторами данных ключевое значение в развитии фазы резистентности при стрессе имеют особенности функции поджелудочной железы и печени. Что касается поджелудочной железы, то это выработка инсулина — важнейшего антистрессового гормона и поддержание его необходимых концентраций на определенном уровне, достаточном для противодействия гормонам стресса. В печени образуются липопротеиды высокой, низкой и очень низкой плотности, которые как показано автором, осуществляют регуляцию функции митохондрий в условиях стрессового воздействия и одновременно доставляют основные субстраты (липиды) к этим энергетическим центрам. Развитие фазы резистентности как раз во многом связано с переключением наработки энергии в тканях за счет преимущественного использования липидов, в процессах прямого окисления их в пероксисомах клеток. Тогда как в фазе тревоги преимущественно используются углеводы в процессах гликолиза и цикла Кребса.
Большое значение в особенностях протекания стресс — реакции имеет чувствительность организма к глюкокортикоидам, воздействующим на рецепторный аппарат клеток- мишеней с перестройкой функции их генетического аппарата и наработкой необходимых для адаптационных процессов белков- ферментов (стуктурный след адаптации). Кроме того глюкокортикоиды усиливают протеолитические процессы в условиях стрессового воздействия. Условно выделяют глюкокортикоид положительные и глюкокортикоид — отрицательные типы, что зависит от наличия или отсутствия в тканях- мишенях глюкокортикоид-рецепторных белково — полипептидных комплексов, подвергающихся в процессе стрессового воздействия протеолизу с образованием аминокислот- адаптогенов [86]. То есть, ГК+ воздействуют при посредстве специфических ГК-рецепторов расположенных на цитоплазматической мембране клетки, в цитозоле и на ее ядерных структурах, а ГК- реагируют непосредственным изменением метаболизма за счет инициации протеолиза глюкокортикоид рецепторными белково – полипептидными комплексами. Точнее у них реагируют все ткани (в большей степени соединительная, лимфоидная, мышечная), но преимущественно на уровне реконструктивного протеолиза, тогда как рецепторный аппарат таких особей менее чувствителен к ГК( Рис.3).
В свою очередь реализация глюкокортикоидного эффекта при стрессовом воздействии зависит от насыщенности ткани определенными аминокислотами и их соотношением. Так, таурин усиливает фунцию и выработку глюкокортикоидов, повышает чувствительность рецепторов клеток к ним [223, 225, 226],в то время как возбуждающие аминокислоты, стимулируют образование АКТГ и выработку катехоламинов в тканях мозга и надпочечников [68, 476]. Следовательно, возбуждающие аминокислоты усиливают в основном первую фазу стресса (адреналовую), а таурин вторую фазу (глюкокортикоидную). А преобладание таурина, либо возбуждающих аминокислот может определять характер гормонального воздействия или тип адаптивного ответа на стрессовую реакцию. Ранее мы приводили сведения о том, что в зависимости от того какие аминокислоты и их производные преобладают в крови можно условно выделить «МОЗГОВЫЕ» и «ПЕЧЕНОЧНЫЕ» адаптивные типы[238].
Имеются данные, что таурин обладает мощным антиоксидантным эффектом, препятствующим образованиям в митохондриях печени свободных радикалов кислорода [481]. И в целом являются достаточно сильным гепатопротектором [144, 214]. В то же время возбуждающие аминокислоты оказывают более выраженный протекторный эффект при различных патологических состояниях на нейроны мозга и в целом на нервную ткань.
Имеются так же сведения, что аргинин повышает выработку контерстрессового гормона инсулина [185].
Имеются так же сведения, что серосодержащие кислоты повышают уровень стрессовых гормонов и устойчивости организма к ним, например, при профессиональных занятиях плаванием [534]. Кроме того, печень под воздействием стрессовых гормонов способствует повышению иммуной системы и устойчивости организма в целом к стрессовому воздействию, кроме того печень под воздействием стрессовых гормонов способствует повышению иммуной системы и устойчивости организма в целом к стрессовому воздействию [473].
К глюкокортикоид — положительным относится большинство млекопитающих, к глюкокортикоид отрицательным- морские свинки, приматы и человек. В печени образуется большое количество белков, которые присутствуют в крови (альбумины, белки острой фазы воспаления и другие полипептидные структуры) подвергающихся катаболизму в условиях стрессового воздействия. Однако у разных организмов, даже одного вида, а тем более у представителей различных видов, этот процесс имеет существенное различие. Таким образом, из имеющихся литературных данных можно сделать предположение, что существует различные адаптивные типы — с преобладанием первой фазы стресса (тревоги) и соответственно гормонов этой фазы и второй фазы- (резистентности) с преобладанием гормонов поджелудочной железы и метаболитов, синтезируемых в печени.
Рис № 3. Схема тетаболических особенностей преобладания в крови аминокислот «мозговиков» и «печеночников» при адаптивных перестройках:
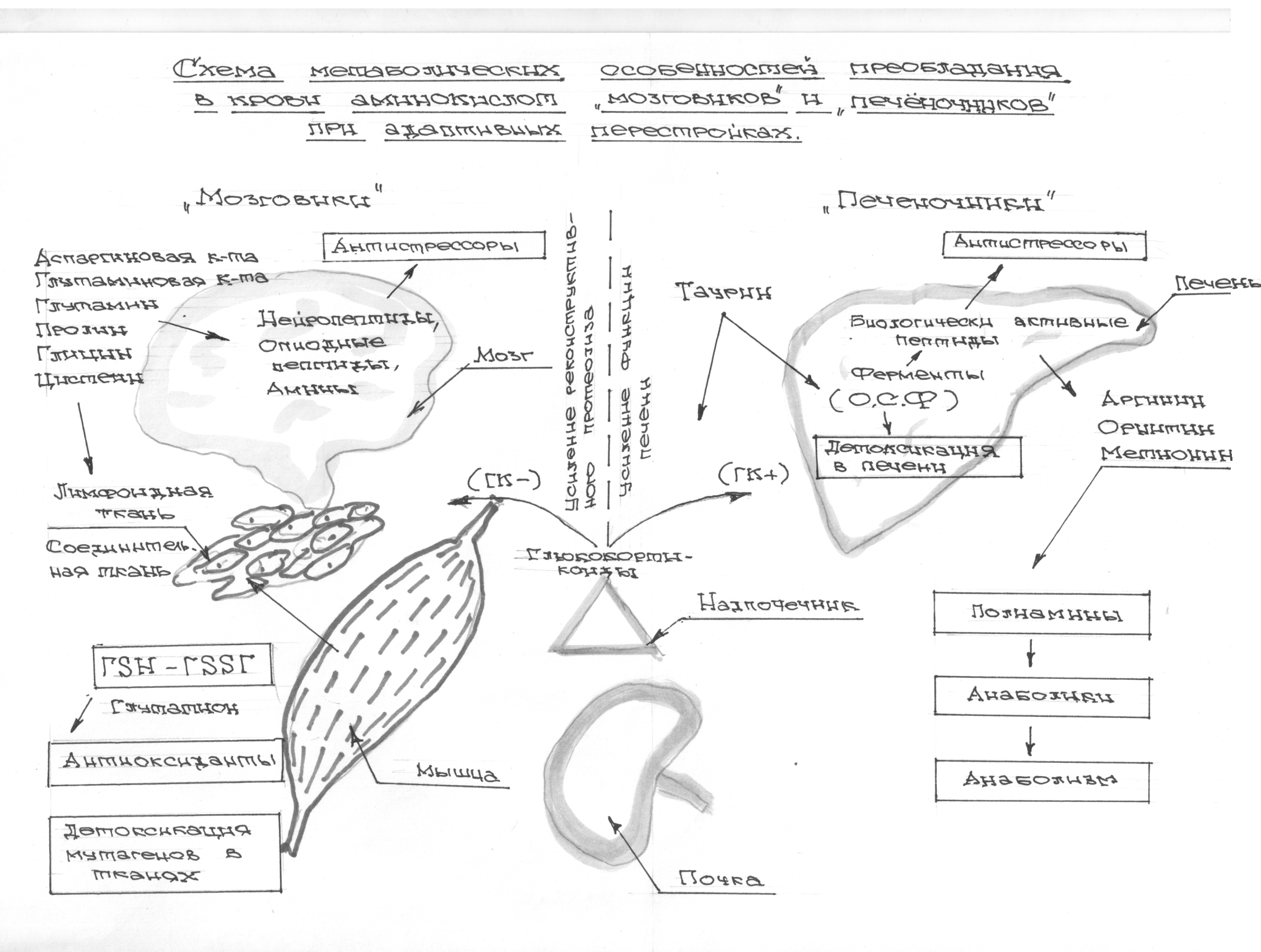
Эти данные в определенной степени были подтверждены в исследованиях Панина Л.Е., Казначеева В.П, Маянского Д.Н.. которые нашли, что как животные, так и люди подразделяются на «тревожных» и «спокойных» особей.
Все это предопределяет разный гомеостатический уровень адаптивных возможностей организма — повышающий или снижающий устойчивость к повреждающим воздействиям.
Важнейшей неспецифической основой изменений метаболизма при стрессе является перестройка окислительно-восстановительных процессов с усилением энергетического обеспечения адаптивных систем в организме. Стрессовое воздействие приводит к нарушению работы основных энергетических центров клеток-митохондрий. За счет повреждения их мембран и ферментных комплексов. Считается, что любое стрессовое воздействие в фазу тревоги сопряжено с генерацией в тканях организма большого количества активных форм кислорода (АФК). Это предопределяется повышенным образованием в хромофинных клетках надпочечников и других отделов нейроэндокринной системы (НЭС) — катехоламинов (КХА), нарушающих нормальное течение окислительно-восстановительных процессов (ОВП) в высокоаэрируемых тканях (легкие, сердце, мозг, кости и др.). Образующиеся при стрессе в больших количествах катехоламины и глюкокортикоиды обезвреживаются в системе оксигеназ смешанных функций (ОСФ) в наибольшем количестве имеющихся в клетках печени, легких, кожи, ряда клеток крови. В процессе обезвреживания образуются промежуточные метаболиты этих веществ, АФК, которые могут повреждать мембранные структуры клеток. В то же время отмеченные процессы способствуют включению компенсаторных механизмов клеток, тканей и систем адаптации. Прежде всего, в тканях усиливается гликолиз, происходит мобилизация и расщепление жира из имеющихся ресурсов организма. Энергетический обмен переключается на использование жира — как более эффективное в условиях повышенной нагрузки и неблагоприятных воздействий на организм. Однако главенствующее значение в условиях повреждающих нагрузок имеет способность организма усиливать свою антиоксидантную защиту (АОЗ) [244].
Необходимо еще раз подчеркнуть, что молекулярные механизмы стресса изучены недостаточно.
В монографии Панина Л.Е. «Биохимические механизмы стресса» 1983 мы находим следующие представления о механизмах стресса.
Известно, что в функционировании нейроэндокринной системы (НЭС) в роли медиаторов выступают КХА, ацетилхолин, серотонин, глутаминовая кислота (и ее производные), глицин, таурин.
На уровне ЦНС вырабатываются пептидные гормоны — рилизинг — факторы, на уровне периферических желез НЭС — гормоны коркового и мозгового слоя надпочечников, В поджелудочной железе — инсулин и глюкагон. Кроме того, в различных тканях и органах вырабатываются тканевые биологически активные вещества — тканевые гормоны: циклические нуклеотиды, простагландины и лейкотриены, а также ряд других метаболитов. На уровне иммунной системы вырабатываются гормоны тимуса, цитокины. Известно, что все эти вещества реализуют свой эффект через рецепторные механизмы. Однако появляется все больше сообщений о том, что их эффект может быть связан с непосредственным включением компонентов этих биологически активных веществ в метаболические процессы.
В последние годы становится все более понятно, что образующиеся в организме свободные радикалы кислорода, азота, а также радикалы обезвреживаемых в тканях веществ экзогенного происхождения (ксенобиотики) и эндогенных метаболитов (эубиотики, например, катехоламины, глюкокортикоиды, производные холестерина, триптофана и ряда других веществ, а также продуктов разрушения мембран клеток, различных белков, липогликопротеидов и т.д.) могут также играть медиаторную функцию — стимуляции, угнетения и сопряжения различных систем организма при повреждающих воздействиях [140,257]. Именно эти промежуточные продукты метаболизма при стрессовых воздействиях могут выполнять роль того загадочного по Г.Селье вещества, которое реализует эффекты стрессового воздействия.
Кроме того, уже много лет, начиная с работ Богомольца, Ужанского известно, что в механизмах аварийного регулирования при гиперстрессе или дистрессе большое значение имеют эндогенные метаболиты — адаптогены — продукты катаболизма, т.е. продукты разрушения тканей и клеточных структур, например, клеток крови (эритродиерез). Они обладают мощным стимулирующим воздействием на защитно-адаптационые системы организма: кроветворную, иммунную, антиоксидантную и другие. Так, мощным стимулирующим, восстанавливающим действием обладают продукты распада клеток при травме, воспалении, гипоксии. Это прежде всего полипептиды, олигопептиды и некоторые аминокислоты: глутаминовая, глутамин, аргинин, пролин, лейцин, валин и ряд других.
Все эти вещества можно объединить как эндогенные адаптогены организма. Однако, несмотря на то, что учение о стрессе существует уже более пятидесяти лет, и многочисленные исследования, посвященные этой проблеме, до сих пор так и не вырисовывается какой-то определенной картины или картин, позволявших бы количественно и качественно охарактеризовать метаболические сдвиги и главные регуляторные перестройки, позволявшие бы эффективно регулировать или вовсе заблокировать нежелательное стрессовое действие или, напротив, создать адекватные модели стрессовых воздействий.
Основная проблема, на наш взгляд, состоит в том, что не известны были те промежуточные фазы регуляции, которые сопрягают неспецифическое звено стресса (ОВП – ПОЛ — АОЗ) и его специфический компонент — реакции иммунного ответа, структурной перестройки на уровне генетического аппарата с последующим образованием на метаболически-генетическом уровне структурного следа адаптации, позволяющего нарабатывать необходимые белки — ферменты и биологически — активные метаболиты для реализации адекватной повреждающему воздействию стресс — реакции.
Мы считаем, что на эту роль могут претендовать аминокислоты и некоторые их производные. Стресс-реакция протекает по-разному у представителей различных ступеней филогенетической лестницы. У человека большое место занимают стрессы, вызванные особенностями жизни и общения между людьми (психоэмоциальные стрессы, воздействие неблагоприятных факторов среды из антропогенных источников — загрязнение биосферы ксенобиотиками, шум, вибрация, заразные заболевания и т.д.), к чему организм человека имеет потенциальные механизмы защиты, начиная от метаболических систем обезвреживания вредных веществ и кончая интеллектом, призванным вырабатывать гармоничные формы существования человека и остальной биосферы.
У животных, стоящих на более низких, чем человек, ступенях филогенетической лестницы стресс-реакция на внешние воздействия менее разнообразна по формам защитных механизмов, участвующих в адаптивных перестройках.
Так, стоящие близко к нам млекопитающие не подвержены таким психоэмоциональным воздействиям как человек, ибо эти воздействия не являются для них адекватными в их жизни.
Как отмечается в монографии Панина Л.Е[244], у пресмыкающихся, земноводных и рыб стресс-реакция по своим механизмам отличается от таковой у млекопитающих.
На определенном этапе филогенеза, разделение первоначально единой нервной системы на вегетативную и соматическую предопределило возникновение трофического аппарата и органов движения.
Нервная система эволюционно связана с эндокринной. Родоначальниками медиаторов нейроэндокринной системы считаются нейрогормоны, такие как катехоламины, эфиры холина, которые обнаружены у бактерий, грибов простейших, растений.
У беспозвоночных уже имеется 2 типа нервной передачи: адренэргический и холинэргический, а также определенную роль в них имеет серотонин.
У низших позвоночных хромаффинная ткань еще не сконцентрирована в симпатических нейтронах и образующиеся в ней адренэргические вещества осуществляют регуляцию гуморальным путем, т.е., через выделение гормонов в кровь.
У высших позвоночных симпатико-адреналовая система заключает в себе два типа клеток — нейроны симпатической нервной системы — (образующие норадреналин) и клетки хромаффинной ткани (образующие адреналин). Важнейшие медиаторы cтpecc-реакции -стероидиые гормоны у рыб и амфибий не разделены на минерал- и глюкокортикоиды. Синтез альдостерна стал возможен только после выхода позвоночных на сушу.
Другим важным гормоном стресс-реакции является инсулин, который как бы противостоит глюкортикоидам и катехоламином в реализации стресса на уровне органов и тканей. Считают, что предшественники В-клеток поджелудочной железы имеются уже у беспозвоночных. У кишечнополостных их обнаружить не удается. У круглоротых и низших позвоночных инсулярный аппарат не сконцентрирован в одном месте и разбросан по печени и кишечнику. У костистых рыб В-клетки концентрируются уже в отдельном органе (брокмановское тельце). Окончательное формирование поджелудочной железы завершается у земноводных и пресмыкающихся.
Нейроэндокринные механизмы, наряду с аллостерическими (активизация или угнетение ферментов, обеспечивающих обменные процессы избытком или недостатком образующегося в этих процессах вещества) составляют фундамент регуляции всех функций организма, предопределяемых метаболическим фоном и создающий метаболический порог адаптивных возможностей организма, представляемых состоянием обмена веществ. Однако тонкая регуляция метаболизма имеет еще более устойчивую основу — своеобразный нулевой цикл, представляемый метаболическим фоном и создающий метаболический порог адаптивных возможностей организма. В зависимости от этого фона происходит реализация регуляторных эффектов гормонов и других регуляторов стресс-реакции.
Этот самый метаболический фон имеет решающее значение при выходе организма из пределов стресс-реакции в исходное состояние или даже повышение резистентности организма к повреждающему фактору и степени обратимости дистресса.
Таким образом можно констатировать, что в процессе филогенеза усложнение адаптивно — функциональных возможностей организма сопряжено с расширением набора биологически активных метаболитов (аминокислот и их производных). В механизмах регуляции адаптивных перестроек — «расширяющаяся метаболическая река филогенеза».
Практически все метаболиты живых клеток, обогащенные азотом: глутамин, аспарагин, аргинин, мочевина, полиамины, нуклеотиды обладают защитными по отношению к кислороду и активным формам кислорода (АФК) свойствами [158]. Эти же вещества стимулируют пролиферацию клеток и рост тканей, т.е., значительное усиление пластических процессов. Это есть своеобразная антиоксидантная защита. Возможно, что, огромная масса тела динозавров именно этим и объясняется. Как мы уже указывали, 60-100 млн. лет назад концентрация кислорода в атмосфере Земли была в 2-3 раза выше, чем сегодня, на чем мы более подробно остановимся ниже.
Однако повышение концентрации 02 в атмосфере за счет пышного и буйного роста растительности сотни миллионов лет назад вследствие интенсивного фотосинтеза приводило помимо появления древнейших животных-гигантов к пожарам в древних лесах, при горении которых образовывалось большое количество ксенобиотиков типа полициклических ароматических углеводородов (ПАУ). Воздействие большого количества 02 и ПАУ могло оказывать мощное мутагенное воздействие на древнейших животных, чем, вероятно, и объясняется их поразительное многообразие. Однако со временем содержание 02 в атмосфере стало снижаться, а ксенобиотиков — нарастать (ввиду их способности накапливаться в биосфере). Появилась проблема защиты организмов от ксенобиотиков и здесь на выручку многоклеточным организмам, очевидно, приходят микроорганизмы, располагающие не только системой цитохромов, использующих 02 в процессах энергетики, но и в процессах пластики и детоксикационных процессах, к которым относится цитохром Р-450 и вся система оксигеназ смешанных функций (ОСФ).
Эта система располагается в цитоплазме клеток многих органов и тканей (более всего в печени, в значительно меньшей степени в легких, кожи, некоторых клетках крови, почках, головном мозге и др.). Она служит для обезвреживания попадающих в организм вредных чужеродных веществ — ксенобиотиков, а также не нужных в организме продуктов распада разрушенных тканей и клеток, излишних биологически активных веществ и продуктов их обмена – эубиотиков.
1.5 Роль трипептида глутатиона и входящих в него аминокислот в механизмах адаптации
Как уже отмечалось выше, нормальное течение окислительно-восстановительных процессов гарантируется достаточно мощной антиоксидантной системой. Это связано с тем, во всех процессах в организме, где используется 02, образуются «побочные» продукты этих реакций – активные формы кислорода (АФК) — супероксидный радикал, синглетный кислород, супероксидный анион, перекись водорода, которые атакуют окружающие мембраны (Владимиров Ю.А., Барабой В.А. 2006, Лукьянова Л.Д.) [65, 29, 189]. В пределах физиологических изменений в организме перечисленные агрессивные вещества обезвреживаются антиоксидантной системой, в которую входят ферменты: супероксиддисмутаза, каталаза, пероксидаза; субстраты — глутатион, витамины А, Е, С, простагландины, глюкокортикоидные, адреналовые гормоны и ряд других веществ. Антиоксидантную систему можно рассматривать как многоступенчатую защиту тканей.
При достаточной мощности антиоксидантной системы обеспечивается необходимая интенсивность окислительных процессов, а, следовательно, и приемлемый в данных условиях уровень адаптивных возможностей организма. Воздействия, превышающие защитные возможности антиоксидантной системы, сопряжены с повреждением клетки, тканей и организма в целом.
Другим важнейшим условием интенсивного протекания окислительных процессов является наличие в достаточном количестве соответствующих субстратов. Субстратами митохондриального и пероксисомального окисления являются производные углеводов, липидов, белков. Субстратами микросомального (для оксигеназ смешанных функций – ОСФ) окисления являются жирорастворимые соединения, поступающие в организм извне, чаще имеющие в своем составе циклические структуры (производные бензола, ПАУ и т.п.), а также эндогенные жирорастворимые вещества: стероидные, адреналовые гормоны, продукты деградации гемоглобина и т.д. Имеются сведения о том, что отсутствие субстратов микросомального окисления значительно увеличивает уровень активных форм кислорода (АФК) в них [227, 85, 83]. Добавление in vitro, например, бенз(а)пирена снижает уровень АФК, следовательно, в роли антиоксидантов могут быть и канцерогенные вещества [148, 149]. Решающее значение в реализации адаптивных возможностей организма имеет не емкость окислительных систем (первая фаза детоксикации), а мощность ферментативных систем второй фазы детоксикации и ее субстратное обеспечение. Увеличение детоксикационных возможностей организма повышает его адаптивные возможности [83,85].
Процесс микросомального окисления рассматривается с позиций его детоксицирующего значения для организма, поскольку вслед за внедрением атомарного кислорода в субстрат в микросомах происходит конъюгация активированных соединений с глутатионом, глюкуроновой кислотой, фосфоаденозинфосфосульфатом, аминокислотами: глицином, глутамином, таурином — и затем выведение из организма [86, 155].
Образующиеся в надпочечниках, щитовидной железе соответствующие гормоны могут играть роль модуляторов описанных выше механизмов.
Ключевую роль в процессах конъюгации занимает глутатион, трипептид, состоящий из глутаминовой кислоты, глицина и цистеина. Одновременно глутатион важен для антиоксидантной системы, обеспечивающей необходимый уровень протекания процессов окисления вообще [83, 85,164, 185]
Продолжительное повреждающее воздействие на организм сопровождается накоплением адреналовых и стероидных стрессовых гормонов, что так же, как и поступление ксенобиотиков может привести к чрезмерной генерации свободных радикалов и истощение субстратов конъюгации, прежде всего глутатиона.
В условиях резкой активации микросомального окисления, усиления (перекисного окисления липидов) ПОЛ, в митохондриях нарушается работа дыхательной цепи, происходит разобщение окисления и фосфорилирования. В этих условиях митохондрии набухают, окислительные процессы в них сопряжены не с образованием энергии АТФ, а с наработкой глутаминовой кислоты [19, 65, 29, 181]. Окислительное фосфорилирование при этом угнетается, стимулируется гликолиз, происходит закисление цитоплазмы и лабилизация мембран лизосом. Последнее определяет появление в клетке активных протеиназ и расщепление белков, что способствует увеличению в тканях свободных аминокислот. Все эти изменения создают условия для стимуляции пролиферативных процессов и одновременно для усиления антиокислительной защиты клетки [126, 200, 394].
Более чем для 50 распространенных патологических состояний доказано ведущее значение в их патогенезе нарушения антиоксидантных возможностей организма [19, 20, 29, 442].
Сюда можно отнести атеросклероз, опухолевый рост, различные воспалительные и инфекционные процессы как бактериальной, так и вирусной этиологии, в том числе туберкулёз.
Показано, что ферментные и субстратные системы, ответственные за защиту клетки от АФК эффективны лишь в интеграции с системой глутатиона. Она является последним и наиболее важным звеном в защите клетки от окислительного стресса [19, 20, 29, 30, 165, 179, 180, 181, 123, 139, 164, 166, 185, 402, 412, 462, 463, 479, 488, 544, 577, 587, 588, 160, 185, 588, 100, 168].
Возможность глутатиона выполнять антиоксидантную и детоксикационную функцию модулируется наличием других антиоксидантных субстратов: витаминов А, С, Е, липоевой кислоты. В эксперименте установлено, что глутатион и альфа-токоферол представляют некую интегральную систему, в которой более стабильное значение имеет первый [83, 397, 487].
Повышенное количество АФК в ткани приводит к быстрому истощению антиоксидантных возможностей альфа-токоферола, тогда как глутатион поддерживается в восстановленном состоянии за счёт работы пентозного цикла и малик-фермента, восстанавливающих НАДФ.
Установлена также тесная взаимосвязь восстанавливающей и антиоксидантной активности глутатиона с обменом аскорбиновой кислоты. Последняя поддерживает глутатион в восстановленном состоянии [170].
Имеется также зависимость антиоксидантной и детоксикационной ёмкости глутатионовой системы от наличия в ткани или инкубационной среде витамина А, который усиливает глутатионпероксидазную активность, а также активность глутатион-S-трансферазы при воздействии на организм ксенобиотиков [83, 85, 168, 227].
При чрезмерной нагрузке свободными радикалами антиоксидантные возможности альфа-токоферола, аскорбиновой кислоты, каротина, простагландинов исчерпывается, и в случае, если система глутатиона угнетается, это сопровождается нарушением биосинтеза белка и повреждением клетки с последующим повреждением мембран, активизацией гидролитических ферментов лизисом, нарушением митохондриального окисления, деградацией метаболизма и гибелью клетки, что имеет место, как уже отмечалось выше, при интенсивном воздействии ксенобиотиков и туберкулезном поражении ткани.
Имея ведущее значение в процессе конъюгации и выведения обезвреживаемых ароматических веществ из организма, глутатион имеет не меньшее значение в механизмах антиоксидантной защиты клетки, нейтрализуя АФК и другие реакционные метаболиты: гидроперекиси липидов, повреждённые компоненты нуклеиновых кислот. И, если в первом случае вступление глутатиона в реакции конъюгации обеспечивается соответствующими глутатион-S-трансферазами, то во втором — глутатион-пероксидазами.
В число ферментов, обеспечивающих конъюгацию выводимых из организма, входят соответствующие трансферазы. Они обеспечивают связывание активированных метаболитов с глутатионом, глюкуроновой кислотой, фосфоаденозинфосфосульфатом и аминокислотами: глицином, глутамином, таурином, цистеином [83, 85, 168, 227].
В экспериментах установлено, что ферменты, обеспечивающие детоксикацию ксенобиотиков, обладают чрезвычайным полиморфизмом. Так, только для цитохрома Р-450 известно несколько десятков изоформ. Это же самое можно сказать о ферментах конъюгации — глутатион-Sтрансферазе, имеющей три изоформы и обладающей к тому же пероксидазной активностью и ответственной за глутатионзависимую защиту от перекисного окисления липидов (ПОЛ), то же самое можно сказать об изоферментах уридиндифосфоглюкуронилтрансферазы. К этому следует добавить чрезвычайный полиморфизм активности ферментов у различных видов животных по отношению к различным субстратам. Имеются видовые отличия в обеспеченности субстратами, необходимыми для конъюгации: глутатионом, глюкуроновой кислотой, аминокислотами.
Введение экспериментальным животным глутатиона или компонентов, из которых состоит этот трипептид: цистеина, глутаминовой кислоты, — частично или полностью предотвращает развитие окислительного стресса под воздействием ксенобиотиков.
Следует подчеркнуть, что защитный эффект отмеченных веществ выражен в большей степени, чем других антиоксидантов. Положительный эффект от введения перечисленных аминокислот был отмечен при различных интоксикациях, воздействии радиации, гипоксии различного генеза.
Накапливается всё больше фактов, что глутатион фокусирует в себе не только антиоксидантные и детоксикационные ресурсы организма: аминокислоты, входящие в этот трипептид, входят в важнейшие метаболические циклы и регуляторные системы различного гомеостатического уровня. Так, биосинтез глутатиона сопряжён с потреблением большого количества 5-оксипролина, источником которого может быть соединительная ткань [170].
Следовательно, обмен глутатиона в определённой степени связан с обменом соединительной ткани, нарабатывающей большие количества пролина и принимающей активное участие в детоксикации ксенобиотиков [300, 301].
Наиболее интенсивно биосинтез глутатиона происходит в печени, откуда он перераспределяется в другие органы. Введение аминокислот-предшественников глутатиона (глутамата и цистеина) интенсифицирует этот процесс. Определенные количества глутатиона образуются и в других органах: лёгкие, почки, лимфоидная, кроветворная ткани и др. Возможно перераспределение глутатиона как между органами, так и между клетками [168, 227].
Глутаминовая кислота, являясь составной частью глутатиона, в большом количестве может образовываться из оксипролина, то есть иметь большие ресурсы в соединительной ткани [68].
То есть, помимо глутатиона в неспецифических механизмах защиты к повреждающему воздействию большое значение имеют аминокислоты, входящие в этот трипептид. Наибольшие изменения в обмене свободных аминокислот при действии неблагоприятных факторов на организм связаны с системой глутатиона. Так, воздействие на крыс пропанола сначала резко изменяет содержание аминокислот в тканях, а затем по мере адаптивных перестроек их уровень нормализуется. Сходные результаты получены в экспериментах по воздействию кровопотери, травматического шока, алкогольной интоксикации.
При экстремальных воздействиях глутаминовая кислота метаболизируются в адаптогены антиоксиданты: ГАМК, ГОМК, янтарную кислоту, аргинин.
Серосодержащие аминокислоты могут быть так же мощными адаптогенами. Цистеин, окисляясь и декарбоксилируясь, превращается в антиоксидант и стабилизатор мембран таурин. При этом метаболиты глутамата, метионина и цистеина реализуют свои антиоксидантные и антистрессорные эффекты как на центральном, так и на периферическом уровне. Эндокринная система мобилизует большое количество катехоламинов и глюкокортикоидов, вследствие чего катаболическим воздействиям подвергаются лимфоидная система и многие белки организма с мобилизацией большого количества аминокислот, влияя на состояние нервной, эндокринной, иммунной систем, периферических тканей и интегрируя таким образом различные уровни адаптивных перестроек.
Следовательно, такие регуляторные метаболиты как аминокислоты — могут являться не только пластическим материалом для биосинтеза различных белков организма, определяющих его специфичность, но и, включаясь в регуляторные и антиоксидантные системы, определять его неспецифическую резистентность.
В нейротрофической регуляции организма принимают активное участие гормоны и другие биологические активные метаболиты. Накапливается все больше фактов, что аминокислоты, то есть, метаболиты, являющиеся родоначальником живой материи, и мобилизующиеся в организме наряду с другими регуляторными метаболитами при повреждающих воздействиях на организм также обладают важным регуляторным воздействием [22, 557, 475, 572, 511, 593, 490, 68, 88, 90, 143, 393, 263, 264, 295, 318, 396, 428, 434, 443, 447, 449, 459, 467, 476, 485, 497, 503, 508, 510, 540, 557, 596, 597, 561, 562, 578].
Так, глутаминовая кислота обладает чрезвычайно широкой биологической активностью и имеет важнейшее значение в адаптации организма к повреждающим воздействиям, прежде всего, сопрягая энергетический обмен с пластическим. В то же время глутамат является иммуномодулятором, стимулирующим лимфоидную систему организма.
Глутаминовая кислота также является нейромедиатором, выполняющим возбуждающую функцию в центральной нервной системе. Но в условиях стрессового воздействия на организм глутамат в ЦНС и других тканях организма интенсивно метаболизируется в гаммааминомасляную кислоту (ГАМК) — мощный тормозный медиатор и антистрессовый метаболит. В процесс е превращения глутамата в ГАМК образуется янтарный полуальдегид, обладающий антиоксидантными свойствами.
Установлено, что применение глутаминовой кислоты и её солей (натриевой и кальциевой) увеличивает концентрацию глутатиона и антиоксидантные возможности организма при высотной, гемической гипоксии, алкогольной интоксикации.
Глутамат нормализует нарушенные функции эндокринной системы (щитовидной железы, надпочечников, поджелудочной железы).
Глутаминовая кислота является фармакопейным препаратом, широко используемым в качестве средства, нормализующего тяжёлые нарушения ЦНС, а также препарата, снижающего повреждающее воздействие противотуберкулёзных препаратов на печень, что также связано с усилением биосинтеза глутатиона.
Кроме того, глутамат оказывает нормализующее воздействие на функцию митохондрий при экстремальных воздействиях на организм, стимулирует биосинтез мочевины при этих воздействиях, способствуя нормализации кислотно-щелочного состояния организма. Глутамат натрия стимулирует костномозговое кроветворение при постгеморрагической анемии. Глутамат натрия модулирует экспрессию цитохрома Р-450 через выработку гормона роста [68, 88, 90].
Изложенное позволяет сделать заключение, что глутаминовая кислота может рассматриваться как адаптоген широкого спектра действия и изменения в её обмене тесно связаны с изменением уровня глутатиона в тканях, а, следовательно, антиоксидантными и детоксикационными ресурсами.
Второй аминокислотой, входящей в глутатион, является глицин, так же, как и глутамат, осуществляющий нейромедиаторную функцию в ЦНС. Глицин также как и глутамат используется при ряде патологических состояний в качестве адаптогена [36, 43, 155, 156, 175].
И, наконец, третьим и последним компонентом глутатиона является цистеин — серосодержащая аминокислота. Цистеин может образовываться только из метионина -незаменимой аминокислоты. Превращение в цистеин возможно лишь после активного включения метионина в процессы метилирования, например, фосфолипидов, креатина, пиримидиновых оснований и других метаболитов. То есть, количество цистеина в тканях зависит от интенсивности включения в обмен метионина. Цистеин обладает антиоксидантной активностью [304, 450, 468, 469].
Глутатионовая конъюгация завершается образованием конъюгатов меркаптуровых кислот, представляющих собой остаток цистеина и процесс обезвреживания эндогенных токсинов и ксенобиотиков сопряжён с выведением из организма большого количества незаменимых серосодержащих аминокислот. Для обеспечения антиоксидантных и детоксикационных процессов должен быть высокий уровень обмена метионина и необходимое поступление его с пищей. В этом плане интересно отметить, что у морских свинок, неспособных синтезировать аскорбиновую кислоту, так же, как и у человека, уровень серосодержащих аминокислот (метионина, цистеина, таурина) в большинстве тканей ниже, а глутатиона, особенно восстановленной его формы, выше, чем у животных, синтезирующих витамин С [236, 237, 239].
Аминокислоты, входящие в глутатион, могут быть предшественниками других мощных антиоксидантных субстратов, нежели глутатион. Так, глутаминовая кислота является мощным стимулятором биосинтеза аргинина и мочевины в печени, являющихся антиоксидантами. Метаболизируясь в орнитин, глутаминовая кислота служит предшественником полиаминов [155, 156, 158]. Аргинин, мочевина и полиамины обладают мощным антиоксидантным потенциалом. Эти вещества накапливаются в тканях при экстремальных воздействиях на организм – например, низкая температура, повышенное давление 02, радиация, физическая нагрузка [158, 133, 70].
Важно отметить, что метионин, активируясь и превращаясь в S-аденозилметионин, включается в процесс биосинтеза полиаминов при резком снижении в тканях глутатиона и увеличении активных форм кислорода (АФК), стимулирующих процессы декарбоксилирования [155, 156, 235].
В первом случае это включение глутамата и серосодержащих аминокислот в глутатион и — во втором — включение их в полиамины, при увеличении биосинтеза аргинина и мочевины в печени, которые стимулируются глутаматом и глутамином. Второй путь преобладает при истощении системы глутатиона и усилении свободнорадикальных процессов в клетке. Именно свободнорадикальному механизму отводится решающая роль в канцерогенном воздействии ксенобиотиков и других неблагоприятных факторов на организм [59, 83, 227, 358]. Возможно, что в данной ситуации организм, исчерпав возможности адекватной физиологическому уровню антиоксидантной и детоксикационной защиты с преобладанием системы глутатиона, переходит на другие защитные механизмы, связанные с образованием большого количества азотистых антиоксидантов: аргинина, мочевины, полиаминов.
Решающее значение в такой перестройке может иметь метаболический фонд свободных аминокислот, прежде всего глутаминовой и серосодеожащих (метионин и цистеин), и уровень активных форм кислорода (АФК) и других радикалов в ткани — влияющих на интенсивность декарбоксилирования.
Представленные механизмы явились основанием для использования глутатиона и серосодержащих аминокислот в качестве веществ, защищающих организм от ксенобиотиков, радиации и ряда других повреждающих воздействий. В то же время при таких воздействиях как повышенное давление кислорода, пониженная температура, защитного эффекта удаётся достичь, используя аргинин, мочевину, то есть, богатые азотом, но не содержащие серу аминокислоты и их производные [159, 158].
Имеются экспериментальные данные о том, что рассматриваемые аминокислоты могут влиять на состояние организма через модуляцию его иммунной системы и ЦНC, интегрируя их функции [334, 410].
Содержание свободных аминокислот и их производных высокостабильно для различных тканей организма. Поэтому изменение этих параметров может существенно изменить метаболический фонд важнейших обменных процессов в клетке.
Мощным источником глутаминовой кислоты и глицина в организме могут быть центральная нервная система (ЦНС) и соединительная ткань (СТ). При болезни Альцгеймера нарушение образования в головном мозге глутамата, глицина и некоторых других аминокислот приводит к нарушениям азотистого обмена во всём организме и его адаптивных возможностей. О том, что значительные количества глутамата и глицина образуются в мозговой ткани, свидетельствуют и другие источники.
Всё изложенное свидетельствует о том, что последствия повреждающих воздействий на организм во многом зависят от его способностей мобилизовать в достаточной степени субстраты, необходимые для эффективного функционирования системы глутатиона, то есть соответствующие аминокислоты.
Имеются многочисленные сведения, что воздействия, предъявляющие повышенные требования к адаптивным системам организма одновременно стимулируют их, что является важнейшим механизмом эволюционного развития [386, 389, 319, 320, 285, 204]. Проведенные нами исследования позволили установить, что различные стресс-воздействия по-разному влияют на различные виды животных, располагающих различными защитно-адаптивными ресурсами, и особенностями специфического и неспецифического ответа на внешние раздражители. Ведущее значение в этих различиях имеют соотношения в тканях и клетках адаптивных систем аминокислот – адаптогенов. В зависимости от ресурса в организме обогащенных азотом или серой аминокислот можно выделить 2 крайних защитно-адаптивных типа – N- и S-тип.
Здесь необходимо дать краткую характеристику процессов совершенствования адаптивных механизмов живой материи по мере эволюционного развития филогенеза.
На сегодня установлено, что жизнь на планете Земля возникла около 3 млрд. лет назад.
Согласно теории Опарина-Холдейна в определенных физико-химических условиях, имевших место на планете в то время, из неорганических веществ (Н2, СО2, NH3, Н2О и ряда других) образовались новые органические молекулы – некоторые аминокислоты, липиды и ряд других.
Зарождение и первые этапы жизни на Земле проходили в анаэробных условиях. Более 2-х млрд. лет господствующей формой жизни на планете были прокариоты – микроорганизмы, во многом предопределившие как основные направления развития живой материи, так и важнейшие процессы геохимии Земли (Шлегель Г. 1987).
Революционное значение в процессах развития живой материи имело появление в водной среде океанов пурпурных зеленых бактерий, а затем цианобактерий – сине-зеленых водорослей, способных к фотосинтезу и образованию О2. Процессы фотосинтеза у бактерий и растений обеспечивали и обеспечивают до сих пор биосферу Земли как органическим материалом так и О2. И то и другое необходимо для непрерывного совершенствования адаптивных механизмов представителей животного мира и приспособления к имеющимся факторам окружающей среды (Шлегель Г, 1981).
Однако уровень О2 в атмосфере, как и структура биомассы Земли, существенно менялись в различные временные периоды. Так, первоначально концентрация О2 была довольно низка – не более 2% вплоть до периода 0,6-0,5 млрд. лет назад. Но по мере того как фотосинтезирующие растения сплошь покрыли все материки концентрация О2 непрерывно возрастала и составила около 100-80 млн. лет назад 60-40% состава атмосферного воздуха (Наука и жизнь, №6 1985).
В образцах воздуха из пузырьков янтаря и древних отложений льда того периода обнаружена именно такая концентрация О2.
Аналогичная концепция о существенном повышении концентрации кислорода во времена каменноугольного периода высказывалась учеными Йельского университета и Американского геологического общества Робертом Бернером и Гарри Линдисом. Значительные концетрации кислорода приводили к интенсивным пожарам и существенно повышали плотность атмосферного воздуха, что делало возможным полет гигантских рептилий и насекомых.
В связи с этим дальнейшая эволюция живой материи потребовала более сложных генетически – метаболических форм и соответственно функционально более совершенных форм организации, нежели прокариоты и растительный мир. Появляются эукариоты – в подавляющем большинстве аэробные организмы. Сначала средой их обитания была водная стихия океанов. Затем они вышли на сушу и распространялись по новым материкам планеты в виде земноводных и пресмыкающихся.
Обращает на себя внимание чрезвычайный полиморфизм и огромная масса последних. Это могло быть связано с несколькими факторами.
300-100 млн. лет назад высокая концентрация О2 в атмосфере неизменно приводила к интенсивным пожарам – сгоранию и перегонке обильно разросшегося растительного мира в результате чего образовывались биологически активные вещества (БАВ) – прежде всего полициклических ароматических углеводородов (ПАУ), накопившиеся в биосфере. (Необходимо отметить, что нефть и уголь, образовавшиеся много миллионов лет назад, содержат большое количество ПАУ). ПАУ – жирорастворимые вещества, легко проникающие через биологические мембраны, резко изменяющие их свойства вплоть до повреждения и разрушения [227, 86].
Центральным звеном обеспечения жизнедеятельности всех живых организмов являются окислительно-восстановительные реакции, но высокие концентрации О2 (сильный окислитель, повреждающий клетки живой материи) и ПАУ поставили взаимодействующую с ними живую материю перед выбором – либо погибнуть или уйти в недоступные для этих веществ ниши – например, глубины океанов, что некоторые представители животного мира и сделали, либо перейти на новый метаболически-адаптивный уровень. И этот переход состоялся, породив новые, гораздо более перспективные в адаптивном плане, чем прокариоты и представители флоры формы жизни – эукариот, представителей фауны.
Развитие животного мира сосредоточилось на совершенствовании аэробных эукариат. – занявших доминирующее положение среди фауны. Использование О2 в качестве окислителя в ОВП резко повысило функционально-адаптивные возможности новых видов и одновременно создало идеальные условия для интенсивного процесса естественного отбора в жестких условиях гипероксидативной среды насыщенной мутагенными БАВ – ПАУ.
С этих позиций становится тоже понятными причины огромного разнообразия обнаруженных ископаемых пресмыкающихся в соответствующие периоды развития биосферы Земли (300- 65 млн. лет назад).
Основываясь на данных современной токсикологии можно попытаться реконструировать адаптивно-метаболические процессы и их совершенствование у животных того времени.
Важнейшее значение в защите важнейших структур клеток и тканей от агрессивных метаболитов при метаболизировании их в тканях являются система детоксикации, протекающая в 2 фазы в эндоплазматической сети клетки. I фаза – окисление проникших в клетку ПАУ в системе ОСФ, чтобы в дальнейшем они могли быть связаны со специальными обезвреживающими веществами и выведены из клеток и тканей организма через почки, легкие, кожу. II фаза – процесс связывания токсичных веществ, в том числе и продуктов обмена (БАВ), с некоторыми специфичными, предназначенными для этого веществами (УДФГК, ФАФС, глутатион, некоторые аминокислоты – глицин, глутамин, таурин). Особенно большое значение в процессах обезвреживания ПАУ и промежуточных продуктов обмена О2 имеет глутатион [227, 86].
Однако в условиях перенасыщенности среды О2 и ПАУ отмеченных механизмов для их обезвреживания недостаточно [158]. Необходимы мощные вещества — адаптогены, защищающие прежде всего от повреждения мембраны. К таким веществам относятся: таурин, полиамины, аргинин, мочевина, нуклеиновые кислоты, мочевая кислота и некоторые другие вещества, образующиеся прежде всего в печени и оказывающие мощное мембраностабилизирующее действие [158, 373, 225]. Одновременно все эти метаболиты оказывают стимулирующий эффект на процессы пролиферации и роста. Возможно, именно эти обстоятельства и определили огромную массу динозавров – как своеобразную защиту к избытку О2 и ПАУ. В то же время высокие мутагенные свойства ПАУ и АФК определяли высокий уровень мутаций, многообразие видов и невысокие адаптивные возможности возникающих видов.
Строго говоря, теория Дарвина, которая связывает естественный отбор с генетическими мутациями, справедлива именно для этого периода развития живой материи. Однако с исчезновением динозавров и появлением новых доминирующих классов животных – млекопитающих значение генетических мутаций в процессах совершенствования защитно-адаптивных механизмов существенно снижается. Как на сегодня признанно большинством специалистов в этих областях научного знания, для млекопитающих большее значение имеет трансгенный перенос, осуществляемый микроорганизмами и размах генетических и фенотипических изменений существенно ниже, чем в предшествующую эпоху [501, 507,].
Поэтому для млекопитающих можно говорить не о генетически-метоболических защитно-адаптивных механизмах, а скорее об адаптивно-метаболических, в которых ведущее значение имеет не значительное изменение генетического аппарата, а изменение метаболических взаимоотношений. В этих модификациях большое значение имеет взаимодействующая с макроорганизмом микрофлора, а не высокие концентрации агрессивных и мутагенных веществ. Поэтому следующим этапом совершенствования адаптивных механизмов после генетически-мутационного можно назвать этапом метаболической компартментализации или метаболически-адаптивный этап.
Снижение концентрации О2 в атмосфере при сохранении в ней большого количества ПАУ (они накапливаются в биосфере и могут долго сохраняться) вследствие массовой гибели реликтовых папоротниковых лесов сделали нежизнеспособными гигантских пресмыкающихся, зависимых от О2 и ПАУ. Доминирующее значение стали приобретать животные, способные эффективно утилизировать О2 и перераспределять его прежде всего для нужд нервной системы – мозга и одновременно способные сохранять устойчивость метаболических процессов и повреждающему воздействию ПАУ. Своеобразным реликтовым видом, обладающим такими возможностями, является крыса (Ж. Сен — Леран 1968).
По мнению многих специалистов именно на крысу по многим параметрам походили первые млекопитающие.
И действительно, особенности обмена веществ у крысы таковы, что они занимают как бы промежуточное положение в плане защитно-адаптивных ресурсов и механизмов между животными, жившими в гипероксидативных условиях и высокой концентрации ПАУ и животными другой эпохи – живущих при гораздо более мягких в токсикологическом плане условиях.
Особенности обмена веществ у крысы состоят в том, что в защитно-адаптивных механизмах отмечается высокая чувствительность к воздействию глюкокортикоидов, мутагенных и канцерогенных веществ, прежде всего ПАУ [368, 369, 86].
Имеются многочисленные работы, в которых показано, что глюкокортикоиды претерпевают те же обменные превращения, что и ПАУ. Продукты их обмена влияют на генетический аппарат клетки, а так же на многие другие рецепторные, мембранные, ферментные и другие регуляторные центры и метаболические процессы [227, 85].
Основой структуры стероидных гормонов, в том числе и ГК, витаминов группы Д, холестерина является молекула циклопентанпергидрофенантрена, то есть ПАУ. Все это наводит на мысль о том, что перечисленные БАВ – это своеобразная интернализация ПАУ — превращение их из экзогормонов среды в эндогормоны организма. В подтверждение этой гипотезы можно привести то обстоятельство, что обезвреживание ПАУ в организме начинается с их окисления при участии цитохрома Р-450 и образование ГК в надпочечниках также происходит с использованием биосинтезирующим аппаратом их клеток цитохрома Р-450 [86].
Реализации эффектов ГК при стресс-воздействии посвящена большая литература. Основным механизмом воздействия считается стимулирование протеолитических ферментов в иммунной, соединительной, мышечной и ряде других тканей организма. Однако многие механизмы ГК воздействия во многом так и не раскрыты, установлено лишь, что имеются некие белково-рецепторные структуры внутри клеток (Rp I, II, III) с которыми связываются ГК. Считается что ГК влияют на генетический аппарат клеток, снимая репрессию со структурных генов и усиливая наработку соответствующих белков-ферментов, кроме того доказано активизирующее влияние ГК на различные ферменты, например тирозин-аминотрансферазу, триптофандиоксигеназу, ферменты глюконеогенеза и ряд других. С другой стороны ГК ингибирует некоторые ферментные системы, например ферменты гликолиза и цикла Кребса.
Несомненным также является тот факт, что ГК, подвергаясь обмену в тканях, способствует генерации большого количества АФК и реакционоспособных метаболитов ПАУ [86].
Повышение количества АФК обеспечивает прямое окисление СЖК при стресс-реакции при повышенном уровне их в организме, с образованием необходимых количеств АТФ для реализации ЗАМ в условиях подавления гликолиза и ЦТК.
Именно при таком типе ответа на стресс-воздействия организм более устойчив к острым и интенсивным повреждающим факторам, что мы и наблюдаем при воздействии таких факторов на крысу.
Необходимо уточнить, что основные детексикационные процессы с участием цитохрома Р-450 по метаболизированию ПАУ, ГК и КХА, а также многих других ксено- и эубиотиков протекают преимущественно в печени.
1.6 МОЗГ И ПЕЧЕНЬ, КАК ДВА ОСНОВНЫХ ПОЛЮСА ОБРАЗОВАНИЯ АДАПТОГЕНОВ ОРГАНИЗМА. «МОЗГОВИКИ» — «ПЕЧЕНОЧНИКИ»
Выдвигаемая нами концепция о различных защитно-адаптивных типах по сути дела является развитием положения о цефализации как о важнейшем механизме эволюционного развития живой материи.
Исходя из наших данных, главенствующее значение в совершенствовании ЗАМ имеет метаболическая функция ЦНС и прежде всего мозга в интеграции с другими защитно-адаптивными системами, прежде всего с печенью, соединительной тканью, системой кроветворения и иммунитета.
При этом ЦНС и мозг играют не только координирующую роль в жизнедеятельности организма, но имеют самостоятельное значение в образовании, накоплении и мобилизации важнейших метаболитов-адаптогенов организма в условиях ответа на стресс-воздействие. Это во многом и предопределяет реализацию и совершенствование ЗАМ и формирование более адаптивных защитно-адаптивных типов.
Особенно большое значение мозг как орган образования адаптогенов имеет при эмоциональном стрессе (ЭС) [22, 263, 264].
Еще раз вкратце напомним, что классические представления о стрессе – это смена фаз: тревоги — резистентности – истощения, во многом предопределяющаяся изменениями гормонально-метаболического фона. Инициируется процесс повышенным образованием в ЦНС и надпочечниках КХА с выбросом их в кровь, стимулирующих ферменты мобилизации из депо углеводов и жиров со значительным повышением их в крови. Это повышение сопровождается усиленным образованием в клетках поджелудочной железы инсулина, обеспечивающего метаболическую утилизацию повышенных количеств углеводов и жиров из крови тканями защитно-адаптивных систем: сердечно-сосудистой, дыхательной, кроветворной и другими, активно участвующими в стресс-реакции. Усиление в тканях ОВП с образованием энергии АТФ позволяет существенно повысить их функциональную активность и сопротивляемость к стресс-воздействию. Однако эти процессы, определяемые воздействием повышенных количеств КХА кратковременны. На смену им идет гормонально-метаболический фон, определяемый образованием повышенных количеств ГК, который собственно и обуславливает основные адаптивные сдвиги в ответ на стрессовый фактор, что было установлено, в основном, на крысах. Прежде всего, происходит резкая активизация лизосомального аппарата клетки и усиление протеолиза белков лимфоидной системе, мышечной и соединительной тканей. Имеются сведения, что ГК (также как и КХА), подвергаясь процессам обезвреживания (при значительном повышении их количества в различных тканях организма — печень, легкие и др.), способствуют повышению уровня АФК и свободных радикалов в форме промежуточных метаболитов обезвреживания КХА и ГК [227, 85, 193, 194, 195]. Это во многом и предопределяет усиление протеолических процессов. Происходит расщепление белков в тканях (прежде всего в соединительной, мышечной) в том числе и многих рецепторных белков как мембранных, так и внутриклеточных (например, рецепторов к ГК – II-III типа) [86], что сопровождается значительным повышением свободных аминокислот в тканях организма. Ряд аминокислот являются мощными адаптогенами. Так, аланин, глутамин, аргинин, аминокислоты с развлетвленной углеродной цепью являются стимуляторами анаболических процессов [158, 296]. Другие аминокислоты обладают антиоксидантным эффектом (глутамат, аргинин, цистеин, таурин). Десять аминокислот стимулируют процессы глюконеогенеза в печени и почках, восполняя тем самым ресурсы углеводов в организме, подвергающимся стресс-воздействию [145]. Все эти механизмы стабилизации метаболических процессов в условиях воздействия повреждающего фактора описываются понятием реконструктивный протеолиз и направлены на выход организма на новый метаболический уровень – более адекватный повреждающему воздействию [251].
Мы считаем, что особенности стресс–реакции и адаптивного ответа на нее определяются различиями на уровне регуляторных метаболических систем, ведущее значение в которых имеют аминокислоты адаптогены. Их ресурс, наследуемые и изменяющиеся в процессе жизнедеятельности организма соотношения.
На то, что стресс-воздействия могут быть разными как по силе, так и по степени и особенностям адаптивного ответа организма указывал еще Г. Селье [293]. Необходимо подчеркнуть, что ЭС существенно отличается от других видов стресса по своим приспособительно-метаболическим механизмам и имеет ключевое значение в механизмах эволюции живой материи вообще и в совершенствовании процессов цефализации в частности. Это особенно важно для становления человека, человеческого общества и его развития. Особенности ЭС состоят в том, что в отличие от других видов стресса, здесь имеет ведущее значение психо-эмоциональный фактор. Основными медиаторами ЭС являются КХА – производные аминокислот фенилаланина и тирозина (норадреналин, адреналин и дофамин) вырабатывающиеся в различных структурах мозга и мозговом веществе надпочечников при I фазе стресса – фазе тревоги [319, 320, 321, 322, 365, 263].
КХА мобилизируют из имеющихся в организме депо углеводы (глюкозу) и липиды (СЖК), способствуя повышенному образованию АТФ в энергетических центрах клеток в процессе их окисления и повышению функциональных возможностей организма, энергетического обеспечения движений – с одной стороны, и для формирования психологических защитных реакций, защищающих от травмирующего психику воздействия (интродукция) [255].
Усиление наработки надпочечниками ГК с реализацией их метаболически адаптивных механизмов при эмоциональном стрессе (особенно при его хроническом воздействии) не имеет столь большого значения. Это происходит потому, что постоянное, многократно повторяющееся психоэмоциональное воздействие обуславливает постоянное образование КХА с реализацией их рецепторно-метаболических механизмов и не дает в полной мере включиться механизмам реализации ГК-эффектов [244, 86].
При воздействии на различных животных ГК и КХА была установлена различная чувствительность к ним различных видов животных. Наиболее значительно отличались по метаболическим последствиям введения ГК крысы и морские свинки. Крысы были высокочувствительны – ГК (+), а морские свинки напротив малочувствительны к ГК (-) [86]. То есть, классические механизмы реализации стресс реакции на метаболическом уровне под воздействием ГК более характерны для крысы, а не для морской свинки. С чем это связано авторы не разъясняют.
Полученные нами результаты позволяют понять, в чем состоят эти различия и более полно представить механизмы адаптивных перестроек в ответ на различные по силе, характеру воздействия повреждающего фактора.
В наших исследованиях установлено, что ГК (+) крысы и ГК (-) морские свинки по-разному реагируют как на ЭС, так и на другие стресс-воздействия. И эти различия связаны, прежде всего, со способностями нарабатывать аминокислоты-адаптогены и некоторые другие метаболиты, обладающие адаптогенным эффектом, в ответ на повреждающие факторы [238].
Одновременно в печени у крыс образуются большие количества таурина, содержание которого повышается в тканях организма, ответственных за повышение его адаптивного уровня. Такой ход метаболических перестроек при более или менее длительном воздействии физического стресс-фактора (температура, радиация, физическая нагрузка, инфекции и т.д.) на организм позволяет легко выйти на метаболический уровень, соответствующей фазе резистентности [237, 238, 239].
В классическом варианте это означает снижение интенсивности образования инсулина поджелудочной железой, причем становится достаточным небольшое увеличение продукции глюкокортикоидов (ГК) надпочечниками для поддержания метаболически-адаптивных механизмов стресса. При этом гормональный фон перестраивается таким образом, что адаптивные ресурсы организма направляются на восстановительно-биосинтетические процессы, стимулируемые на уровне генетического аппарата глюкокортикоидами по механизму дерепрессии генов-операторов. Это определяет усиление работы структурных генов, передающих информацию на информационную РНК, с последующим образованием на рибосомах клетки необходимых для адаптационных перестроек белков-ферментов, с усилением сопротивляемости к повреждающему воздействию. Это мы и наблюдали у крыс при воздействии на них различных видов гипоксии, токсичных доз ксенобиотиков, кровопотери, заражения МБТ, проникающей реакции. Эти же воздействия на морских свинок приводили к довольно скорой их гибели, без существенных адаптивных перестроек, но они были высокоустойчивы к мутагенным воздействиям, а также к эмоциональному стрессу. Что определяло столь существенные различия?
В экспериментах на животных нами было установлено, что многие метаболические процессы, имеющие отношение к адаптивным перестройкам у этих видов животных были различными [237,238,239]. Так, у морских свинок в тканях было в 2 раза больше железа и серы, чем у крыс. Основное количество железа и серы в тканях входит в железосерные комплексы окислительно-восстановительных ферментативных центров, определяющих интенсивность протекания окислительно-восстановительных процессов (ОВП) в митохондриях (ферменты дыхательной цепи) и оксигенез смешанных функций в цитозоле клеток [361,378]. В митохондриях железосерные центры ферментов необходимы для процессов окислительного фосфорилирования и образования энергии АТФ. В цитозоле клетки ферменты с железосерными комплексами необходимы для детоксикации ксенобиотиков либо токсичных веществ, образующихся в ходе некоторых промежуточных метаболических превращений собственных веществ (эубиотиков), подвергающихся разрушению в системе оксигеназы смешанных функций (ОСФ), включающих в себя цитохром Р-450.
Имеются сведения, что у крыс в большинстве тканей, особенно в печени намного больше цитохрома Р-450 и связанных с ним ферментов ОСФ, чем цитохромооксидаз или других ферментов дыхательной цепи митохондрий, необходимых для процессов окислительного фосфорилирования и включающих в себя железо и серу [227].
Но исходя из наших данных при сниженных по сравнению с крысой детоксикационных возможностях железа и серы в тканях морских свинок намного больше, чем у крыс, следовательно, оно может находиться преимущественно в железосерных комплексах митохондрий, а не в ОСФ. Или другими словами морские свинки – это «митохондровики» ориентированные на процессы Эмбдена-Меергофа-Кребса. Тогда как крыс можно характеризовать как «ОСФ-ников – пентозников». Это становится еще более понятно, если учесть, что в условиях стресса повышенный ГК фон тормозит процессы расщепления и окисления углеводов (вследствие угнетения образования инсулина), а также процессы окислительно-восстановительного фосфорилирования с их разобщением. Энергия АТФ при этом образуется за счет прямого окисления жирных кислот, за счет стимуляции их перекисного окисления высоким фоном активных форм кислорода (АФК) в митохондриях [86]. В этих же условиях стимуляция глюконеогенеза повышает уровень образования глюкозы из аминокислот, способствует стимуляции гексозомонофосфатного шунта (ГМФШ) и наработки НАДФН2, необходимого для интенсивного протекания ОСФ, прежде всего в печени, а также образованию аскорбиновой кислоты в соединительной ткани. Кроме того, усиленное образование НАДФ Н2 изменяет процессы обмена и функцию мембран, а также функции многих систем, напрямую зависящих от состояния мембран, например, гепатоцитов в печени или нейронов в мозге [170]. Соответственно меняется их функционально-метаболические возможности.
Кардинальное отличие в метаболизме у этих животных состоит в том, что морские свинки (также как приматы и человек) не могут синтезировать аскорбиновую кислоту (АК). Поэтому стресс-воздействие у крыс сопряжено с усиленным образованием АК и изменением обмена соединительной ткани (СТ) в сторону повышенного образования оксипролина и усиленным образованием СТ с потреблением необходимых для этого аминокислот – пролина (глутамата) и глицина с уменьшением их количества в крови [237].
1.7. ЭКСПЕРИМЕНТАЛЬНЫЕ ИССЛЕДОВАНИЯ
1.7.1. РОЛЬ ОСОБЕННОСТЕЙ МЕТАБОЛИЗМА АМИНОКИСЛОТ СОЕДИНИТЕЛЬНОЙ ТКАНИ В МЕХАНИЗМАХ АДАПТАЦИИ ОРГАНИЗМА К ОТРИЦАТЕЛЬНЫМ ФАКТОРАМ ОКРУЖАЮЩЕЙ СРЕДЫ УСТАНОВЛЕННЫХ В ОПЫТАХ НА МОРСКИХ СВИНКАХ И КРЫСАХ
Животные имеющие различное строение соединительной ткани по -разному реагируют на отрицательные факторы окружающей среды. Крысы, имеющие грубоволокнистую соединительную ткань (СТ), устойчивы к заражению микобактериями туберкулёза (МБТ) и токсическому воздействию ксенобиотиков. Морские свинки — животные, имеющие нежноволокнистую и рыхлую СТ, высокочувствительны к повреждающим воздействиям этих факторов, но они высокоустойчивы к мутагенным и канцерогенным воздействиям. Ключевое значение в отмеченных различиях имеет преобладание в защитных метаболических механизмах аскорбиновой кислоты (АК), таурина либо системы глутатиона с интенсивным окислением этого трипептида. АК, таурин препятствуют окислению глутатиона и повреждающим эффектам молекул биорегуляторов – активных форм (АФК) и высокоактивных соединений азота (ВСА). Специфические особенности обмена аминокислот определяют неспецифические регуляторные воздействия данных биологически активных молекул.
Рыхлая СТ может быть своеобразным ресурсом аминокислот (глутамат, цистеин, глицин) для биосинтеза глутатиона у морских свинок, а грубоволкнистая у крыс своеобразным детоксикатором и антиоксидантом.
Выявленные механизмы могут свидетельствовать о том, что нарастающие в последние годы явления дисплазии соединительной ткани (ДСТ) могут быть связаны с усиливающимся воздействием мутагенных факторов, при снижении устойчивости организма к инфекции, токсичным и другим острым воздействиям. То есть такая стратегия адаптации связана с избеганием интенсивных повреждающих воздействий и защитой хромосомного аппарата от хронических малоинтенсивных воздействий (мутагенов, психокоммуникативных стрессов и т.д.).
В последние годы все более актуальной для медицины становится проблема широкого распространения такого явления как дисплазия соединительной ткани (ДСТ) [1, 38, 8, 377]. Явления ДСТ сопровождаются самым разнообразными патологическими процессами — от нарушений психики до тяжелых расстройств кровообращения. Соединительная ткань имеет тесные взаимосвязи с нервной, кроветворной, иммунной системами, системой детоксикации и другими адаптивными системами организма. Однако в чем причины возникновения дисплазии соединительной ткани (ДСТ) и какое значение она может иметь в изменении адаптивных механизмов организма к меняющимся условиям среды во многом не известно [8, 136, 233, 246, 301]. Поэтому целью наших исследований было выявить взаимосвязь метаболических процессов, определяющих специфику адаптивного ответа в организме подопытных животных на воздействие широко распространенных неблагоприятных факторов окружающей среды (ксенобиотиков и микроорганизмов), с обменом соединительной ткани (СТ).
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Для экспериментов были взяты 2 группы животных – крысы и морские свинки – соединительная ткань которых, как и адаптивно – метаболические процессы резко отличаются. Так СТ крыс грубоволокнистая и плотная, а у морских свинок рыхлая и нежноволокнистая [259]. Морские свинки, в отличие от крыс, не способны синтезировать аскорбиновую кислоту (АК) имеющую большое значение в обмене СТ. Возможно именно поэтому они высоко чувствительны к такому широко распространенному отрицательному фактору окружающей среды как микобактерии туберкулеза (МБТ) [247]. В то же время морские свинки высокоустойчивы к мутагенным и канцерогенным воздействиям, а крысы несмотря на высокую устойчивость к МБТ легко поддаются мутагенному и канцерогенному воздействию в эксперименте [168, 358, 359, 368, 369, 580]. В эксперимент было взято по 10 морских свинок и крыс в контрольных группах и по 10 в опытных. Всего 40 морских свинок и 40 беспородных белых крыс питомника Уральского НИИ фтизиопульмонологии.
В наших исследованиях животные подвергались заражению МБТ в дозе 0,1 мг живой культуры в хвостовую вену – крысы (10 животных) и в дозе 0,001мг в паховую складку – морские свинки живой культуры микобактерий штамм H37Rv (10 животных) и интратрахеальному введению мутагенных и канцерогенных ПАУ: (морские свинки — 10 животных и крысы – 10 животных) адсорбированных на саже в виде масляного раствора каменноугольной смолы (КС) [368, 125].
Определялось мутагенное воздействие КС в микроядерном тесте [229].
Исследовалось количество оксипролина в тканях, уровень аскорбиновой кислоты и ее производных [115, 125].
Кроме того сравнивалось токсическое воздействие больших доз КС на морских свинок (10 животных) и крыс (10 животных) – по 2 грамма ежедневно на кожу.
У всех зараженных животных исследовались параметры экспериментального туберкулеза: морфологические данные, микробиологические исследования, индексы поражения и весовые коэффициенты [247].
Методом ионообменной хроматографии на аминокислотном анализаторе ААА339 «Микротехника» определялось количество свободных аминокислот и их производных в замороженных в жидком азоте тканях исследуемых животных. На основании полученных данных исследовалась динамика количества аминокислот, связанных с обменом СТ (пролин, оксипролн, глутамат), генерацией активных форм азота и образованием перекисей липидов и аминокислот, участвующих в биосинтезе глутатиона (аргинин, цистеин, глицин), а также содержание глутатиона восстановленного и окисленного.
Активные формы кислорода (АФК) и высокоактивные соединения азота (ВСА), являясь высокоактивными биологическими молекулами, оказывают регуляторное воздействие на обмен СТ, усиливая катаболические процессы в ней [136,246,530,589]. Поэтому с помощью иммуноферментного анализатора определялся уровень производных оксида азота и продуктов перекисей липидов в крови исследуемых животных. При этом использовавли следующие диагностические тест системы (определение перекисей липидов по OXISTAT- BiomedicaGRUPPE, определение стабильных производных оксида азота-RandDSystem)
Животные выводились из эксперимента через 2 недели после заражения МБТ — в период, когда у морских свинок развивался генерализованный туберкулез и через 3 месяца после запыления КС. Токсическое воздействие оценивалось по гибели большей части в животных в какой -либо группе.
Экспериментальные исследования были проведены в Уральском НИИ фтизиопульмонологии. Выражаем глубокую благодарность всем сотрудникам, помогавшим в работе.
ПОЛУЧЕННЫЕ РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Заражение МБТ
После заражения МБТ через 2 недели у морских свинок развивается генерализованный туберкулез. У крыс даже при использовании гораздо большей дозы МБТ специфический процесс не развивается. Некоторые органы морских свинок при этом значительно увеличиваются. Так масса печени возрастает с 20,9 до 34,4г, селезенки с 1,06 до 4,3г, легких с 3,8 до 9,6г. Отмеченное увеличение происходит за счет интенсивного включения пролина в оксипролин соединительной ткани, количество которого, например, в печени, возрастает на порядок. Так количество оксипролина в ткани печени у интактных морских свинок составляет 88,2мг/100г сухой ткани, а после заражения его количество возрастает до 650,6мг/100г, то есть в 7 раз (р< 0,01). В крови зараженных морских свинок при этом количество пролина снижается в 2,5 раза ( р< 0,01). Очевидно, он используется для биосинтеза оксипролина в печени.
Изменения массы органов крыс после заражения не происходит, хотя количество оксипролина в печени также несколько возрастает, хотя статистически достоверного отличия не наблюдается (р >0,05).
При воздействии каменноугольной смолы КС каких -либо нарушений в костном мозге морских свинок не отмечено, при том что у крыс количество микроядер в костном мозге увеличивается в 3 раза (р <0,01).
При интенсивном токсическом воздействии КС (2г. на кожу) через 2 недели погибают все морские свинки, тогда как крысы все остаются живы.
При отмеченных явлениях резко изменяется и приобретает диаметрально противоположный характер обмен аскорбиновой кислоты (АК) в тканях сравниваемых животных.
В печени крыс после заражения значительно возрастает количество АК (почти в 3 раза — р <0,01), а у морских свинок (которые в отличии от крыс не могут ее синтезировать) наблюдается ее более чем 2 кратное уменьшение (р<0,01) (Табл.1.1)
Табл 1.1 Содержание аскорбиновой кислоты в печени морских свинок и крыс после заражения МБТ (в мкг/г) (M+_ m(min-max)):
| Группы животных (кол-во в группе) | Контроль (№10) | Заражение МБТ(310) |
| Морские свинки | 142,4+ 21,5
(83,0 – 196,7) |
74,2+ 15,4**
(25,3 — 123,1) |
| Крысы | 252,6+ 42,4
(132,7- 369,5) |
736,3+ 112,5**
(284,3- 1376,2) |
Значком**- отмечено статистически достоверное отличие (р<0,01)
Противоположные изменения претерпевают и метаболиты аскорбиновой кислоты (АК). В печени морских свинок резко увеличивается содержание дикетогулоновой кислоты (ДКГК) (448,6+- 62,4 мкг/г до 864,2+-67,5мкг/г- (р<0,01)) а у крыс наоборот отмечается тенденция к уменьшению ее количества (186,4 +-16,5мкг/г до 215,8мкг/г – (р >0,05)).
Противоположную направленность имеют изменения количества такого метаболита как глутатион, имеющего тесную взаимосвязь с обменом АК.
У морских свинок в печени в 3 раза (р<0,01) прирастает количество глутатиона окисленного (Г-SS-Г) которое и в контроле было несколько больше, чем у крыс. Надо отметить, что и ресурс глутатиона восстановленного (Г- SH) в печени морских cвинок гораздо выше, чем у крыс (712,5мкг/г против 433,2мкм/г- (р<0,01)) (Табл 1.2, Рис. 1.1).
Табл 1.2 Содержание глутатиона восстановленного и окисленного в ткани печени морских свинок и крыс после заражения МБТ (в мкг/г сырого веса ткани) (M+_m(min-max):
| Морские свинки | Контроль (интактные) (№10) | Заражение МБТ(№10) |
| Глутатион восстановленный | 712,5+87,5
(462,7- 1158,0) |
585,7+59,4
(349,6- 822,9) |
| Глутатион окисленный | 465,9+72,8
(246,8- 725,0) |
1438,8+218,1
(869,3- 2150,9)** |
| Крысы | Контроль (интактные) (№10) | Заражение МБТ(№10) |
| Глутатион восстановленный | 433,2+31,5
(366,1- 465,8) |
296,7+58,9
(112,5- 420,2) |
| Глутатион окисленный | 394,6+89,6
(197,5- 771,4) |
356,2+96,5
(97,8 – 577,2) |
Значком* отмечены результаты статистически достоверно отличающиеся от контроля (р<0,05); **- (р<0,01).
Значком* отмечены результаты статистически достоверно отличающиеся от контроля (р<0,05); **- (р<0,01).
Рис. 1.1:
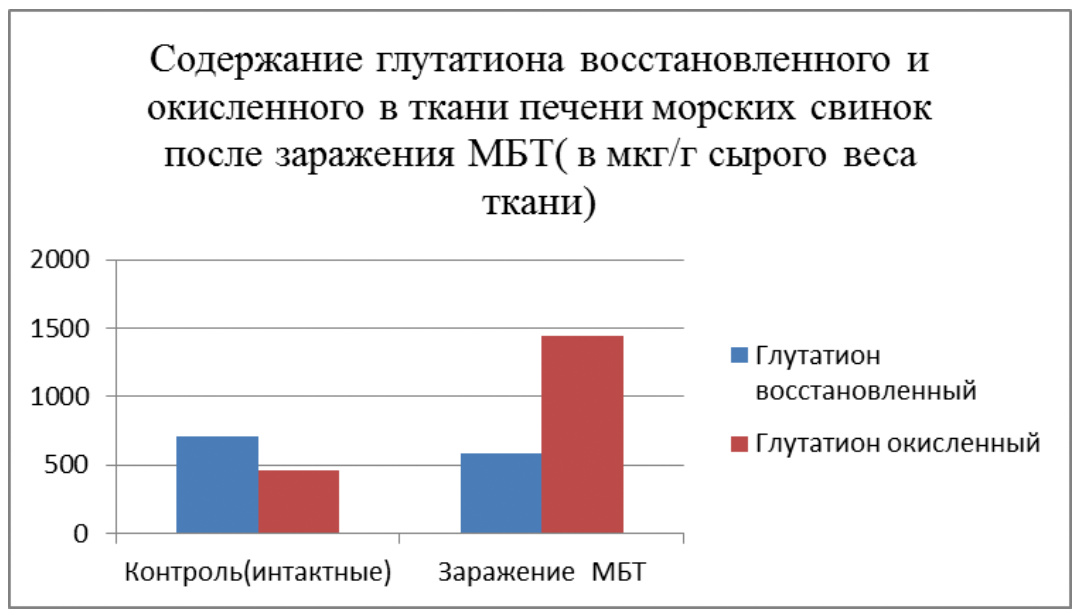
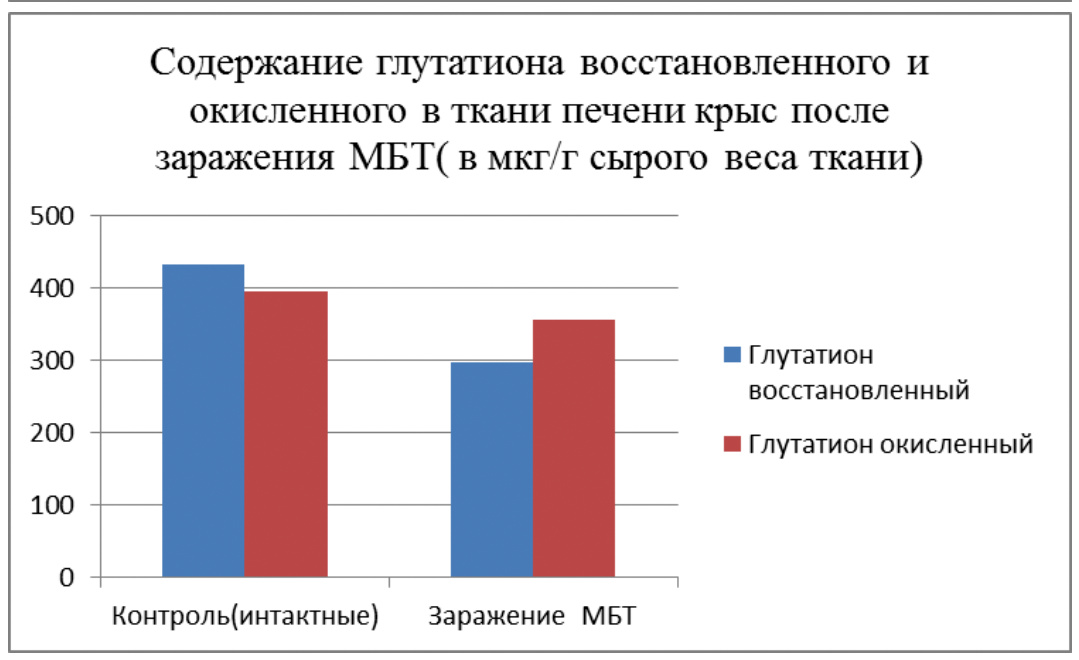
Как видим в защитных механизмах к воздействию МБТ у крыс преобладает аскорбиновая кислота (АК) и усиливается ее биосинтез. А у морских свинок содержание АК резко уменьшается, но увеличивается ее метаболит ДКГК, который может включаться в метаболические процессы необходимые для поддержания глутатиона в восстановленном состоянии (пентозный цикл). При этом общее количество глутатиона у морских свинок резко возрастает и он интенсивно окисляется. АК и глутатион мощные антиоксиданты и биологические молекулы выполняющие важнейшую защитно – адаптивную функцию на метаболическом уровне [65, 74, 238, 450, 452, 487]. Как видим преобладание АК у крыс на этом уровне связано с их высокой устойчивостью к МБТ, а преобладание глутатионовой системы у морских свинок делает их абсолютно беззащитными к заражению туберкулезом.
При этом установлено, что помимо АК в высокой устойчивости крыс к МБТ может иметь такой метаболит как таурин, являющийся антиоксидантом, стабилизатором мембран и стимулятором фагоцитарных реакций организма [223, 224, 238, 281, 281, 595]. Среди всех свободных аминокислот тканей крыс таурин составляет до 50% и более. А у морских свинок таурина в тканях на порядок, а то и на 2 меньше, чем у крыс. Так в костном мозге крыс содержание таурина составляет 5859,1+620,4мкм/л, а у морских свинок 468,7+71,1 мкм/л), почти столь же значительны отличия и по содержанию глутатиона восстановленного в костном мозге (Г-SH) – 422,9+67,1мкм/л у морских свинок против 1199,4+142,5 у крыс (p<0,01).
Значительны отличия и по другим аминокислотам. Так, ранее мы публиковали данные о том, что в тканях крыс, устойчивых к МТБ, особенно в лейкоцитах и кроветворных органах, преобладают такие аминокислоты как таурин, аргинин, метионин, а в тканях морских свинок (особенно в печени) преобладают глутамат, цистеин, глицин – то есть метаболиты необходимые для биосинтеза глутатиона [237, 238, 239, 281, 282].
Обращает на себя внимание, что при этом в крови морских свинок отчетливо возрастает уровень перекисей липидов и высоко активных соединений азота. То есть наростает уровень АФК и ВСА – важнейших регуляторных биомолекул способных выполнять как защитные, так и повреждающие функции [59, 65, 84, 121, 123, 130, 131, 140, 141, 258, 289, 290, 450, 452, 487, 547]. В контроле у морских свинок количество производного ВСА — нитрата в крови превышает таковой у крыс (р<0.05) (Табл. 1.3). После заражения морских свинок уровень ВСА в крови прирастает еще примерно на 50%, а у крыс не изменяется. Уровень перекисей липидов (по оксистату) в крови морских свинок после заражения МБТ взрастает почти в 2 раза (р<0,01), а у крыс статистически значимо на изменяется. Надо сказать, что в контроле количество перекисей у морских свинок в 2 раза выше, чем у крыс (р<0,01).
Табл 1.3 Содержание стабильных производных оксида азота (нитрата) и суммарных перекисей липидов (по оксистату) в крови морских свинок после заражения МБТ (M+_m(min-max):
| Группы животных (количество в группе) | ||
| NO3 (мкмоль/л) | Oxistat (мкмоль/л) | |
| Интактные(№10) | 56,3+11,2
( 43,2-83,7) |
394 +126,1
(29,7- 938,1) |
| Заражение МБТ(№10) | 88,8+10,5
(54,0- 207,3)* |
609,5+143,4
(135,2- 1374,8)* |
Табл 1.4 Содержание производных оксида азота (нитрат) и суммарных перекисей липидов (по оксистату) в крови крыс зараженных МБТ(M+_m(min-max):
| Группы животных
(количество в группах) |
NO3 (мкмоль/л) | Oxistat(мкмоль/л) |
| Интактные (№10) | 26,1+2,9
(18,7- 34,3) |
145,0 + 31,6
(25,7- 185,2) |
| Заражение МБТ (№10) | З1,9 + 4,4
(15,7- 39,7) |
163,1 + 38,8
(27,6-278,5) |
Значком* отмечены результаты статистически достоверно отличающиеся от контроля (р<0, 05), **( р<0,01).
Такое различие в уровнях метаболитов АФК (активных форм кислорода) и ВСА (высокоактивных соединений азота) после заражения МБТ очевидно связано с различием в стратегии защитных механизмов у сравниваемых животных. Так в печени у крыс после заражения более чем в 3 раза прирастает количество туарина (до 1657,5 мкг/г), которое и без того превосходило таковое у морских свинок в 4 раза (р<0,01) (Табл. 1.4).
В то же время у морских свинок в печени столь же значительно прирастает количество глутатиона окисленного (Г-SS-Г) (до 1438,8 мкм/г), которое и в контроле было несколько больше, чем у крыс (Табл. 1.2). Надо отметить, что и ресурс глутатиона восстановленного (Г- SH) в печени морских cвинок гораздо выше, чем у крыс (712,5 против 433,2мкм/г).
Табл.1.5 Содержание свободных аминокислот и их производных, связанных с биосинтезом мембран в печени морских свинок и крыс в мкг/г сырой массы печени в контроле и после заражения МБТ(M+_m(min-max):
| Морские свинки | Контроль(№10) | Заражение МБТ (№10) |
| фосфоэтаноламин | 87,4 +17,3 | 299,7+28,4 |
| (33,2 – 96,6) | (213,6- 430,8)** | |
| этаноламин | 71,3+7,4 | 103,7+9,8 |
| (44,5- 95,1) | (71,2+-145,4)* | |
| Серин | 161,8+17,3 | 199,8 +18,9 |
| (98,4-220,6) | (99,0- 268,1) | |
| таурин | 129,8+1 7,6
(84,4- 232,6) |
251,6+31,4
(157,8- 418,3)* |
| Крысы | Контроль(№10) | Заражение
МБТ(№10) |
| фосфоэтаноламин | 212,1+54,6
(126,7- 328,4) |
«0»- «следы»)** |
| этаноламин | «следы» | «0» |
| Серин | 182,4+28,5
(119,- 306,2) |
84,5+ 26,9
(39,2- 176,4)** |
| таурин | 525,2+28,9
(440,6- 583,2) |
1657,5+425,4
(813,2- 3816,7)** |
Значком* отмечены результаты статистически достоверно отличающиеся от контроля (р<0,05); **- (р<0,01).
Кроме того, как уже было отмечено в печени крыс после заражения значительно возрастает количество аскорбиновой кислоты (АК) (почти в 3 раза), а у морских свинок (которые в отличии от крыс не могут ее синтезировать) наблюдается ее более чем 2 кратное уменьшение (Табл.1.1). Мощный антиоксидантный эффект АК и таурина, естественно, не позволяют чрезмерно увеличивать уровень активных форм кислорода (АФК) и высокоактивных соединений азота (ВСА) в тканях крыс после экстремальных воздействий и ограничивают их повреждающее воздействие. Так в наших исследованиях установлено, что после заражения МБТ в печени крыс исчезают такие метаболиты как фосфоэтаноламин (ФЭА), этаноламин (ЭА) и серин — метаболиты необходимые для биосинтеза мембран (Табл. 1.4). То есть происходит усиление пластических процессов с повышением стабилизации мембран клеток печени, необходимое для усиления детоксикационных, анаболических и других адаптивных механизмов.
У морских свинок возможности такой стабилизации нет. В чем же смысл высокого уровня АФК и ВСА в тканях морских свинок при экстремальных воздействиях? Или это специфично только для туберкулезного процесса?
Несмотря на то, что морские свинки практически беззащитны даже к единичным вирулентным МБТ, но они высокоустойчивы к мутагенным и канцерогенным воздействиям. Вызвать у них экспериментальный опухолевый процесс очень трудно [358, 359, 368, 369].
Воздействие каменноугольной смолы (КС)
Итак, стратегия метаболической адаптации морских свинок к воздействию МБТ ориентирована на усиленное образование глутатиона с его интенсивным включением в окислительные процессы. Что абсолютно не защищает морскую свинку по отношению к МБТ, но может быть чрезвычайно важно для защиты генетического аппарата от мутагенных и канцерогенных воздействий. Человек, приматы и морская свинка не способны синтезировать аскорбиновую кислоту (АК) но зато резистентны к мутагенам и канцерогенам. В механизмах защиты при этом ведущее значение имеет система глутатиона [65, 68, 123, 129, 131, 168, 358, 359, 547].
Табл 1.6 Содержание глутатиона восстановленного и окисленного в ткани печени морских свинок и крыс после воздействия каменноугольной смолой (в мкг/г сырого веса ткани)(M+_m(min-max)):
| Морские свинки (№8) | Контроль (№10)
(интактные) |
Воздействие КС (№10) |
| Глутатион восстановленный | 712,5+87,5
(462,7- 1158,0) |
260,2+18,3
(171,7- 293,3)** |
| Глутатион окисленный | 465,9+72,8
(246,8- 725,0) |
1291,4+ 118,9
( 974,2- 1671,8)** |
| Крысы (№8) | Контроль (№10)
(интактные) |
Воздействие КС (№10) |
| Глутатион восстановленный | 433,2+31,5
(366,1- 465,8) |
363,6+ 31,5
(294,6- 460,3) |
| Глутатион окисленный | 394,6+89,6
(197,5- 771,4) |
557,6+106,1
(341,8- 954,3) |
Значком* отмечены результаты статистически достоверно отличающиеся от контроля (р<0,05); **- р<0,01.
Введение КС почти в 3 раза увеличивает количество глутатиона окисленного и снижает количество глутатиона восстановленного в печени морских свинок(p<0,01). У крыс таких изменений количества глутатиона не происходит (Табл.1.6). При этом воздействие КС в 3 раза (p<0,01) повышает количество микроядер в костном мозге крыс, то есть проявляет отчетливый мутагенный эффект. У морских свинок такого влияния каменноугольной смолы (КС) не наблюдается.
Такая стратегия адаптации у морской свинки сопряжена с понижением устойчивости к МБТ и многим другим вирулентным бактериям (очевидно в связи с особенностями ее жизни на островах Южной Америки). Не высока устойчивость морских свинок и к токсическим воздействиям. Мегадозы КС убивают их в течение 1 – 2 недель, тогда как крысы все остаются живы.
Воздействие каменноугольной смолы (КС) сопровождается повышением уровня ФЭА (фосфоэтаноламина), ЭА (этаноламина) и серина в печени морских свинок (Табл. 1.7). То есть наблюдается повреждение и лабилизация мемран. Все это происходит на фоне значительного повышения уровня перекисей липидов и высокоактивных соединений азота в крови морских свинок. Высокий уровень Г-SS-Г, АФК и ВСА усиливают апоптоз клеток [85, 131, 140, 141, 257, 258], что так же, как и повышенный ресурс глутатиона и его функциональная активность является механизмом антимутагенеза и антиканцерогенеза [83, 85, 168, 227]. Такие механизмы блокируются у крыс большим количеством таурина и АК в тканях и отчасти объясняет легкость моделирования на них мутагенеза и канцерогенеза.
В наших предшествующих исследованиях установлено, что применение таурина повышает устойчивость морских свинок к МБТ и эффективность лечения туберкулеза (с более быстрой нормализацией количества лейкоцитов, моноцитов и лимфоцитов крови) у больных туберкулезом легких [281, 282]. При этом снижается уровень АФК и ВСА в крови больных туберкулезом легких людей и экспериментальных животных. В лейкоцитах подопытных морских свинок увеличивается количество таурина, глутатиона, аргинина.
Более быстрое стихание острофазовых воспалительных реакций и нормализация иммунных нарушений наблюдалась нами при применении глутамата натрия у больных инфильтративным туберкулезом [238, 239]
Табл.1.7 Содержание свободных аминокислот и их производных, связанных с биосинтезом мембран в печени морских свинок и крыс (в мкг/г сырой массы печени) в контроле и после воздействия каменноугольной смолы (КС)(M+_m(min-max)):
| Морские свинки (количество в группе) | Контроль (№10) | Воздействие КС (№10) |
| Фосфоэтаноламин | 87,4 +17,3
(33,2 – 96,6) |
188,5+ 33,1
(108,3-485,7)** |
| этаноламин | 71,3+7,4
(44,5- 95,1) |
«0»** |
| Серин | 161,8+17,3
(98,4- 220,6) |
«0»** |
| таурин | 129,8+17,6
(84,4-232,6) |
162,9+28,1
(60,8- 247,6) |
| Крысы
(количество в группе) |
Контроль (№10) | Воздействие КС (№10) |
| фосфоэтаноламин | 212,1+-54,6
(145,2- 316,8) |
«0»** |
| этаноламин | «следы» | «0» |
| Серин | 182,4+ 28,5
(119,- 306,2) |
85,2+12,6
(54,7- 116,2)** |
| таурин | 525,2+28,9
(440,6- 583,2) |
1124,5+ 86,2
(886,3- 1366,4)** |
Значком* отмечены результаты статистически достоверно отличающиеся от контроля р <0,05; ** — р <0,01.
А вот введение аргинина и его предшественника глутамата усугубляет течение экспериментального туберкулеза, видимо, за счет усиления образования NO (аргинин является субстратом для образования NO) и его повреждающего эффекта [238, 239, 281, 282].
В то же время имеются данные, что аргинин стимулирует нейроэндокринную и иммунную системы экспериментальных животных [524, 526]. То есть многое зависит от того каких животных используют.
Заражение крыс МБТ, особенно на фоне введения им аргинина и его предшественника глутамата, уменьшает мутагенное воздействие КС на крыс [239]. В крови таких животных резко повышается уровень АФК и ВСА и резко нарастает уровень глутатиона окисленного в печени до 1588,9+138,9 (общее количество глутатиона восстановленного и окисленного становится 1688,3 – то есть значительно больше, чем в контроле и при изолированном воздействии этих факторов), но количество микроядер в костном мозге уменьшается в 2 раза (p<0,05). То есть для этого более важно усиление окисляемости глутатиона (как в и тканях у морских свинок) а не наличие его восстановленной формы, высоких концентраций таурина, аргинина и АК (как в тканях у крыс). Возрастает общее количество глутатиона в костном мозге крыс при совместном воздействии КС и МБТ. Концентрация таурина при этом резко падает.
ВЫВОДЫ:
1. Различное строение соединительной ткани у морских свинок и крыс сопряжено с преобладанием в защитных механизмах к воздействию МБТ и КС глутатиона или аскорбиновой кислоты и таурина.
2. При преобладании в защитных механизмах системы глутатиона (как у морских свинок) с интенсивным его окислением организм более устойчив к мутагенным воздействиям при усилении образования оксида азота в организме.
3. При преобладании в защитных механизмах аскорбиновой кислоты и таурина (как у крыс) организм более устойчив к заражению МБТ при большей устойчивости мембран клеток к АФК и ВСА, но при этом менее устойчив к мутагенному воздействию КС
4. Защитные либо повреждающие эффекты АФК и ВСА неспецифичны и зависят от стратегии защитных механизмов в организме, в которых может преобладать система глутатиона, либо таурин, блокирующий систему глутатиона и его окисление.
5. Таурин блокирует окисление глутатиона (очевидно его эффект стимуляции апоаптоза) и препятствует его антимутагенному действию с повышением количества ВСА.
6. МБТ снижает количество таурина в организме и способствует увеличению роли системы глутатиона в антимутагенных эффектах, такое воздействие стимулируется глутаматом и аргинином – субстратом для образования NO.
7. Нарастающие в последние годы явления ДСТ могут быть связаны с усиливающимся мутагенным и канцерогенным воздействием на человека с преобладанием в защитных механизмах глутатиона и обеспечением его биосинтеза необходимыми ресурсами из соединительной ткани (глутамат, глицин, цистеин), при снижении роли в защитных механизмах детоксикационной роли печени (биосинтез таурина).
1.7.2. РОЛЬ ОСОБЕННОСТЕЙ МЕТАБОЛИЗМА АИНОКИСЛОТ И ИХ ПРОИЗВОДНЫХ В ПОСТРОЕНИИ ФОСФОЛИПИДОВ МЕМБРАН У ЖИВОТНЫХ С РАЗЛИЧНОЙ ЧУВСТВИТЕЛЬНОСТЬЮ К ТУБЕРКУЛЕЗУ И МУТАГЕННЫМ КСЕНОБИОТИКАМ (МОРСКИЕ СВИНКИ И КРЫСЫ)
Актуальность проблем, связанных с распространением туберкулезной инфекции и повышения резистентности к ней не уменьшается. Одним из факторов снижения устойчивости организма к МБТ может быть воздействие ксенобиотиков нарушающих защитные возможности организма. К таким воздействиям можно отнести мембранотропные мутагенные и канцерогенные ксенобиотики продукты перегонки органического топлива (ПАУ) — широко распространенные и накапливающиеся в окружающей среде. От состояния мембран фагоцитов во многом зависит полноценность фагоцитоз (завершенность фагоцитоза) по отношению к внедрившимся в организм возбудителям туберкулеза.
Целью исследования было уточнить особенности обмена фосфолипидов мембран и факторов, влияющих на них у животных с перекрестной чувствительностью к МБТ и мутагенным и канцерогенным ксенобиотикам — у морских свинок и крыс. Что могло бы помочь выявить факторы естественной резистентности к туберкулезу и воздействию мутагенных ксенобиотиков.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Морские свинки восприимчивы к заражению туберкулезом в то же время они высокоустойчивы к мутагенным и канцерогенным воздействиям, а крысы несмотря на высокую устойчивость к МБТ легко поддаются мутагенному и канцерогенному воздействию в эксперименте [237, 238, 358, 359].
В эксперимент было взято по 10 морских свинок и крыс в контрольных группах и по 10 в опытных. Всего 40 морских свинок и 40 беспородных белых крыс питомника Уральского НИИ фтизиопульмонологии.
В наших исследованиях животные подвергались заражению МБТ в дозе 0,1 мг живой культуры в хвостовую вену – крысы (10 животных) и в дозе 0,001мг в паховую складку – морские свинки живой культуры микобактерий штамм H37Rv (10 животных) и интратрахеальному введению мутагенных и канцерогенных ПАУ: (морские свинки — 10 животных и крысы – 10 животных) адсорбированных на саже в виде масляного раствора каменноугольной смолы (КС) [368].
Определялось мутагенное воздействие КС в микроядерном тесте [229].
Кроме того сравнивалось токсическое воздействие больших доз КС на морских свинок (10 животных) и крыс (10 животных) – по 2 грамма ежедневно на кожу.
У всех зараженных животных исследовались параметры экспериментального туберкулеза: морфологические данные, микробиологические исследования, индексы поражения и весовые коэффициенты.
Методом ионообменной хроматографии на аминокислотном анализаторе ААА 339 «Микротехника» определялось количество свободных аминокислот в замороженных в жидком азоте тканях исследуемых животных. На основании полученных данных исследовалась динамика количества аминокислот связанных с обменом фосфолипидов мембран (фосфоэтаноламин (ФЭА), этаноламин (ЭА), серин, таурин, метионин, глутатион и участвующих в биосинтезе глутатиона (глутамат, цистеин, глицин), аргинина, а так же содержание глутатиона восстановленного (Г-SН) и окисленного (Г-SS-Г).
Активные формы кислорода (АФК) и высокоактивные соединения азота (ВСА), являясь высокоактивными биологическими молекулами, оказывают воздействие прежде всего на мембраны клеток и тканей. Развитие туберкулезного процесса, так же как и воздействие мембранотропных циклических ксенобиотиков сопряжено с генерацией АФК и ВСА опосредующих повреждающее воздействие на мембраны клеток [140, 141, 289, 290, 257, 258]. Поэтому с помощью иммуноферментного анализатора определялся уровень производных оксида азота и перекисей липидов в крови исследуемых животных. При этом использовавли следующие диагностические тест системы (определение перекисей липидов по OXISTAT- BiomedicaGRUPPE, определение стабильных производных оксида азота-RandDSystem).
Животные выводились из эксперимента через 2 недели после заражения МБТ — в период, когда у морских свинок развивался генерализованный туберкулез и через 3 месяца после запыления КС. Токсическое воздействие КС оценивалось по гибели большей части в животных в какой-либо группе.
ПОЛУЧЕННЫЕ РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
На момент выведения из эксперимента у всех зараженных морских свинок развивался генерализованный туберкулез с преимущественным поражением селезенки, печени, лимфоузлов, легких. У крыс, не смотря на то что доза заражения была в 100 раз большей, туберкулезный процесс не развивался.
Установлено, что у животных с различной видовой устойчивостью к МБТ генерация высокоактивных соединений азота (ВСА) в организме в ответ на заражение музейным штаммом МБТ -Н37 Rv значительно отличается. Так у морских свинок количество конечного продукта окисления ВСА – нитрата в плазме крови возрастает после заражения от 56, +11,2 в контроле до 88,8 +10,5 мкмоль/л, то есть примерно в 1,6 раза (р < 0,01 к контролю). Тогда как у крыс достоверного увеличения этого вещества к контролю не наблюдается (26,14 – 31,9 мкмоль/л соответственно). Следует отметить, что морские свинки располагают вообще большими ресурсами ВСА так как в контроле содержание нитрата в их плазме более чем в 2 раза превосходит таковой у крыс.
Обращает на себя внимание, что у морских свинок так же гораздо более высокие показатели суммарных перекисей липидов определяемых по OXISTAT. Их содержание в крови у здоровых морских свинок 394,46+54,8 мкмоль/л., у зараженных — 609,53+143,40. У крыс его содержание 145,4+31,6 – 163,8+18,8 мкмоль/л соответственно, то есть, изменений после заражения практически не наблюдается. Разница между морскими свинками и крысами по показателям суммарных перекисей липидов в контрольных группах примерно в 2 раза.
Но после заражения МБТ это различие становится почти 4х кратным. Причем у крыс увеличения показателей по OXISTAT практически не наблюдается, а у морских свинок его уровень резко возрастает (почти в 2 раза). Нитрат является конечным метаболитом высокоактивных соединений азота (ВСА) генерируемых эндотелием, фагоцитами, а показатели по OXISTAT — регистрация конечных метаболитов превращения перекисей липидов (ПОЛ) под воздействием активных форм кислорода (АФК) [168, 257]. Поэтому динамика значений нитрата и ПОЛ в крови является неким интегральным показателем способности организма генерировать ВСА и АФК.
Защитный или повреждающий эффект АФК и ВСА при туберкулезе зависит от состояния антиоксидантной и других защитных систем организма [258, 452].
Такое различие в уровнях метаболитов АФК И ВСА после заражения МБТ очевидно связано с различием в стратегии защитных механизмов у сравниваемых животных. Так в печени у крыс после заражения более чем в 3 раза прирастает количество таурина (до 1657,5 мкг/г), которое и без того превосходило таковое у морских свинок в 4 раза (р<0,01) (Табл. 1.5).
После заражения МБТ у морских свинок в 3 раза увеличивается количество фосфоэтаноламина (ФЭА), в 1,5 раза — этаноламина (ЭА) и имеется тенденция к увеличению количества серина. Скорее всего, это следствие усиления катаболических процессов при развитии туберкулезного воспаления (Табл. №5).
Но после воздействия каменноугольной смолы (КС) в печени у морских свинок примерно в 2 раза повышается уровень ФЭА при том, что уровень ЭА и серина резко падает (до «0» значений) (Табл.1.7). То есть биосинтез фосфолипидов скорее всего усиливается, но ресурс их непосредственного предшественника ФЭА сохраняется на повышенном уровне.
У крыс при воздействии МБТ и КС наблюдаются однотипные сдвиги в содержании предшественников фосфолипидов: резкое уменьшение количества ФЭА и ЭА (до «0» значений) и примерно в 2 раза уменьшается количество серина. То есть наблюдается интенсивное потребление этих метаболитов для биосинтеза фосфолипидов мембран и с усилением анаболических процессов с истощением метаболитов необходимых для биосинтеза фосфолипидов (Табл. 1.5, 1.7). Характерно, что при этом существенно возрастает количество метионина и аргинина (стимуляторов мноцитарного ростка кроветворения и фагоцитоза [450] в костном мозге. От «следовых» значений до 51 мкг/г и 24,3 мкг/г соответственно. Количество аргинина при этом возрастает в 2- 3 раза (Р <0,01 в обоих случаях). Но при этом в кроветворной ткани крыс под воздействием МБТ и КС в 2-3 раза уменьшается содержание Г-SН и Г-SS-Г, соответственно, во столько же сокращается общая сумма глутатиона (Табл.1.2, 1.6).
Скорее всего анаболический и мощный мембраностабилизирующий эффект на состояние мембран оказывает таурин в большом количестве присутствующий в печени крыс, которое еще более прирастает после воздействий микобактериями туберкулеза и каменноугольной смолой (МБТ и КС). Таурин является антиоксидантом, стабилизатором мембран, средством детоксикации [456]. С чем очевидно и связан гораздо более низкий уровень АФК и ВСА в крови крыс по сравнению с морскими свинками и их более высокая выживаемость при интенсивном воздействии КС.
Среди всех свободных аминокислот тканей крыс таурин составляет до 50% и более, что в несколько раз, а иногда и на порядок, превосходит таковое в тканях морских свинок.
Значительны отличия и по другим аминокислотам. Так ранее мы публиковали данные о том, что в тканях крыс устойчивых к МТБ, особенно в лейкоцитах и кроветворных органах, преобладают такие аминокислоты как таурин, аргинин, метионин, а в тканях морских свинок (особенно в печени) преобладают глутамат, цистеин, глицин – то есть метаболиты необходимые для биосинтеза глутатиона [238, 452].
После заражения МБТ в печени крыс исчезают такие метаболиты как фосфоэтаноламин (ФЭА), этаноламин (ЭА) и серин — метаболиты необходимые для биосинтеза мембран. То есть происходит усиление пластических процессов с повышением стабилизации мембран клеток печени, необходимое для усиления детоксикационных, анаболических и других адаптивных механизмов.
У морских свинок возможности такой стабилизации нет. В чем же смысл высокого уровня АФК и ВСА в тканях морских свинок при экстремальных воздействиях? Или это специфично только для туберкулезного процесса?
Несмотря на то, что морские свинки практически беззащитны даже к единичным вирулентным МБТ, но они высокоустойчивы к мутагенным и канцерогенным воздействиям. Вызвать у них экспериментальный опухолевый процесс очень трудно [100, 359].
При токсическом воздействии КС (2г. на кожу) через 2 недели погибают все морские свинки, тогда как крысы все остаются живы.
Однако при менее интенсивном, но более длительном воздействии КС у крыс количество микроядер в костном мозге увеличивается в 3 раза (р<0,01), то есть проявляется мутагенный эффект на кроветворную ткань. В то время как у морских свинок мутагенного воздействия при этом не отмечалось. Скорее всего, это было связано с гораздо более мощной и активной, чем у крыс системой глутатиона — важнейшего антимутагенного и антиканцерогенного метаболита при воздействии мембранотропных ксенобиотиков [100, 168]. У морских свинок после воздействия МБТ или КС количество Г-SН и Г-SS-Г и соответственно их суммы взрастает в кроветворной ткани в 2- 3 раза (P<0,01) (Табл.№2,6), что, очевидно, является следствием его биосинтеза в печени и перераспределения в кроветворную ткань.
После заражения МБТ у морских свинок в печени в 3 раза (р<0,01) прирастает количество Г-SS-Г, которое и в контроле было несколько больше, чем у крыс. Надо отметить, что и ресурс Г- SH в печени морских cвинок гораздо выше, чем у крыс (712,5мкг/г против 433,2мкм/г- р<0,01) (Табл. 1.2).
Итак, стратегия метаболической адаптации морских свинок к воздействию МБТ ориентирована на усиленное образование глутатиона с его интенсивным включением в окислительные процессы. Что абсолютно не защищает морскую свинку по отношению к МБТ, но может быть чрезвычайно важно для защиты генетического аппарата от мутагенных и канцерогенных воздействий. Человек, приматы и морская свинка не способны синтезировать аскорбиновую кислоту, но зато резистентны к мутагенам и канцерогенам. В механизмах защиты при этом ведущее значение имеет система глутатиона [100, 168, 359].
В печени морских свинок после воздействий каменноугольной смолой (КС) количество Г-SS-Г увеличивается примерно в 3 раза и почти во столько же снижается концентрация Г-SH (Табл.1.6). Но суммарное количество глутатиона при этом увеличивается. То есть усиливается его биосинтез, что видимо и предотвращает мутагенное воздействие КС на морских свинок.
У крыс существенного увеличения общего количества глутатиона после воздействий КС не отмечается, равно как и его окисленной формы. То есть система глутатиона активно не включается в антиоксидантные и детоксикационные механизмы, что возможно связано с блокированием ее таурином.
Такая стратегия адаптации у морской свинки сопряжена с понижением устойчивости к МБТ и многим другим вирулентным бактериям (очевидно в связи с особенностями ее жизни на островах Южной Америки). Не высока, как видим, устойчивость морских свинок и к токсическим воздействиям.
Воздействие каменноугольной смолой сопровождается повышением уровня ФЭА, ЭА и серина в печени морских свинок. То есть наблюдается повреждение и лабилизация мембран. Все это происходит на фоне значительного повышения АФК и ВСА в крови морских свинок. Высокий уровень Г-SS-Г и АФК и ВСА усиливают апоптоз клеток, что так же как и повышенный ресурс глутатиона и его функциональная активность является механизмом антимутагенеза и антиканцерогенеза [100, 358, 359]. Такие механизмы блокируются у крыс большим количеством таурина в тканях, что отчасти и объясняет легкость моделирования на них мутагенеза и канцерогенеза.
В наших предшествующих исследованиях установлено, что применение таурина повышает устойчивость морских свинок к МБТ и эффективность лечения туберкулеза (с более быстрой нормализацией количества лейкоцитов, моноцитов и лимфоцитов крови) у больных туберкулезом легких [238]. При этом снижается уровень АФК и ВСА в крови больных туберкулезом легких людей и экспериментальных животных. В лейкоцитах подопытных морских свинок увеличивается количество таурина, глутатиона, аргинина.
Более быстрое стихание острофазовых воспалительных реакций и нормализация иммунных нарушений наблюдалась нами при применении глутамата натрия у больных инфильтративным туберкулезом [238].
А вот введение аргинина и его предшественника глутамата усугубляет течение экспериментального туберкулеза, видимо, за счет усиления образования NO (аргинин является субстратом для образования NO) и его повреждающего эффекта [238, 257, 258].
В то же время имеются данные, что аргинин стимулирует нейроэндокринную и иммунную системы экспериментальных животных [456]. То есть многое зависит от того каких животных используют.
Ранее мы публиковали сведения о том, что заражение крыс МБТ, особенно на фоне введения им аргинина и его предшественника глутамата, уменьшает мутагенное воздействие КС на крыс [238]. В крови таких животных резко повышается уровень АФК и ВСА и нарастает уровень глутатиона с его интенсивным окислением, но количество микроядер в костном мозге при этом уменьшается в 2 раза (p<0,05). То есть для этого более важно усиление окисляемости глутатиона (как в и тканях у морских свинок) а не наличие его восстановленной формы, высоких концентраций таурина, аргинина и аскорбиновой кислоты (как в тканях у крыс). Возрастает общее количество глутатиона в костном мозге крыс при совместном воздействии КС и МБТ. Концентрация таурина при этом резко падает.
ВЫВОДЫ:
- Метаболизм предшественников фосфолипидов мембран у морских свинок и крыс под воздействием МБТ и КС имеет свои особенности.
- У крыс при обоих воздействиях изменения однотипны и характеризуются резким снижением количества ФЭА, ЭА, серина, что может быть связано с усилением анаболических, антиоксидантных, мембраностабилизирующих, детоксикационных механизмов.
- У морских свинок изменения разнонаправлены. При воздействии МБТ значительно повышается уровень ФЭА, ЭА, серина, что может отражать усиление катаболических, прооксидантных механизмов вследствие резкого повышения уровня АФК и ВСА, снижения детоксикационных процессов. При воздействии КС становится неопределяемым уровень ЭА и серина, тогда как уровень ФЭА повышается, что может отражать усиление биосинтеза фосфолипидов при одновременном повышении ресурсов для их образования.
- На особенности метаболизма фосфолипидов мембран при рассматриваемых воздействиях у морских свинок и крыс влияют различия в обеспечении этих животных регуляторными серосодержащими метаболитами.
- У крыс в печени при обоих воздействиях значительно увеличивается содержание таурина — мощного антиоксиданта, анаболика, стабилизатора мембран. Очевидно, это позволяет мобилизовать из печени метионин и аргинин (стимуляторов моноцитарного ростка кроветворения и фагоцитоза) количество которых резко возрастает в костном мозге. Но при этом в несколько раз снижается уровень глутатиона. Это видимо обеспечивает устойчивость крыс к МБТ, но не спасает их от мутагенного воздействия КС на кроветворную ткань.
- У морских свинок при воздействии МБТ и КС в печени и костном мозге в несколько раз возрастает уровень глутатиона и его окисляемость. Это не защищает морскую свинку от МБТ, но предотвращает мутагенное воздействие КС.
- Учитывая полученные результаты естественная резистентность к МБТ очевидно может быть усилена таурином, метионином, аргинином, ФЭА, ЭА, серином, а к мутагенному воздействию ПАУ- ксенобиотиков – глутатионом и аминокислотами в него входящими, ЭА, серином.
- Таким образом метаболический стереотип при таких экстремальных воздействиях как заражение подопытных животных туберкулезом и воздействии токсичных и мутагенных ПАУ (каменноугольная смола) определяет особенности функционирования мембран, а те в свою очередь определяют характер адаптивного ответам на стрессовое воздействие, во многом обеспеченного определенными аминокислотами- адаптогенами (в нашей терминологии по типу «мозговики» — «печеночники»).
1.8 АМИНОКИСЛОТЫ И ИХ ПРОИЗВОДНЫЕ ОПРЕДЕЛЯЮЩИЕ «МОЗГОВЫЕ» и «ПЕЧЕНОЧНЫЕ» АДАПТИВНЫЕ ТИПЫ ПРИ СТРЕССОВЫХ ВОЗДЕЙСТВИЯХ
Таким образом из всего представленного материала следует, что как среди животных, так и среди людей можно выделить преимущественно «МОЗГОВЫЕ», так и «ПЕЧЕНОЧНЫЕ» адаптивные типы. В защитно-адаптивных механизмах, которых на метаболическом уровне преобладает либо мозг, либо печень. Печень во многом обеспечивает функционально-метаболические изменения в организме в реализации стресс-воздействия и адаптивного ответа на него. И от ее ресурсов во многом зависит тип адаптивного ответа на стресс-воздействие.
Именно печень обеспечивает организм многими адаптогенами в условиях стресс-воздействия и адаптации крыс. В печени мобилизуются многие аминокислоты-адаптогены, транспортные белки и пептиды, серосодержащие аминокислоты, основные ресурсы Fe, Cu, Mn и других микроэлементов, здесь образуются транспортные формы липидов (ЛПОН, ЛПНП, ЛПВП) используемые в условиях стресса как энергетические ресурсы, а также как регуляторные факторы энергетических процессов в условиях стресс-воздействия. Кроме того, печень во многом ответственна за стабилизацию температурного режима организма, особенно в условиях холодового стресса [244].
Здесь усиливается наработка НАДФН2 в ГМФШ и образование аскорбиновой кислоты. Все это повышает адаптивные резервы организма, прежде всего его окислительно-восстановительные возможности и обеспечивает резистентность крыс к таким повреждающим факторам, как токсические дозы ПАУ, кровопотеря, высотная гипоксия, радиация, но не к хроническому эмоциональному стрессу, мутагенным и канцерогенным воздействиям.
У млекопитающих печень составляет около 4% от массы тела – это самый большой паренхиматозный орган, обеспечивающий адаптивные метаболические перестройки. Однако у приматов и человека печень составляет всего 2% от массы тела, примерно столько же, как и головной мозг (2%) [378].
Мозг также является крупнейшим паренхиматозным органом, способным нарабатывать мозговые адаптогены организма [22]. Так, здесь образуются и в больших количествах присутствуют адаптогены широкого спектра действия, защищающие ткани организма при повреждающих воздействиях: глутаминовая кислота, аспарагиновая кислота, глутамин, ГАМК, ГОМК, цистотионин, цистеиновая кислота, таурин, изотеиновая кислота. В мозге образуются и другие БАВ, обладающие мощными адаптивными эффектами: различные гаммаглутамилпептиды (например, глутатион), катехоламины, серотонин, эндорфины, энкефалины, олиго- и полипептиды, обладающие адаптогенным эффектом в условиях стресс-воздействия. Некоторые из этих веществ выполняют медиаторные функции в различных отделах мозга [262, 263, 365, 268, 322, 327, 328]. Для других какая-либо определенная функция в головном мозге не установлена (карнозин, анзерин, цистатионин, изотеиновая кислота, N-ацетилглутаминовая кислота, N-ацетиласпарагиновая и др.) В то же время эти вещества обладают антиоксидантными, мембраностабилизирующими свойствами, например, в сердечной мышце, скелетной мускулатуре и других органах и тканях [197, 199]. Таким образом, мозг является не только органом, воспринимающим и обрабатывающим сигналы из внешней среды и затем корректирующий организм сообразно изменяющейся ситуации, но и наряду с печенью являющийся мощным органом метаболической адаптации, нарабатывая БАВ-адаптогены, способные переходить через геметоэнцефалитический барьер (ГЭБ) и участвовать в стабилизации адаптивных систем организма [158, 263, 22, 224, 225].
В то же время между мозгом и печенью имеется взаимодействие при реализации стресс-воздействия. Так, вещество, обладающее общим тормозным воздействием на возбудимые ткани – таурин, образуется преимущественно в печени, но имеет специальные транспортные системы в ГЭБ и мозговых структурах, концентрируется в тканях мозга, особенно в его сером веществе, существенно повышая тормозные ресурсы нейронов [158, 263, 223, 224, 22].
С другой стороны, глутаминовая кислота и глутамин в больших количествах образуются в мозге, особенно в условиях стресс-воздействия. В условиях стресса в тканях мозга освобождается большое количество аммиака, очень токсичного для нервной ткани. Связывание и, соответственно, обезвреживание аммиака осуществляется глутаминовой кислотой в глутаминсинтетазной реакции с образованием больших количеств глутамина. Глутамин легко проникает через ГЭБ и используется прежде всего в печени в биосинтетических процессах [22]. Глутамат и глутамин используются в различных адаптивно-метаболических процессах организма прежде всего в печени и почках [235, 68, 155, 239].
Однако характер реализации стресс-воздействия может быть различным. Так, крысы и морские свинки существенно отличаются по чувствительности к воздействию стресс-факторов. Другие млекопитающие находятся в промежутке между 2-мя этими видами по чувствительности к ГК в ходе реализации стресс-реакции и адаптивного ответа организма.
Высокий уровень АФК в тканях у крыс стимулирует декарбоксилазы цистеиновой кислоты и орнитина и соответственно образование таких адаптогенов как таурин, полиамины прежде всего в печени и некоторых других тканях организма (Oguro T 1987) [523].
Высокий уровень АФК способствует окислению АК и НАДФН2 и стимулирует тем самым ГМФШ, а усиление глюконеогенеза и реконструктивного протеолиза и значительное повышение количества в тканях свободных аминокислот обеспечивает его необходимым субстратом – глюкозой. Эти же обстоятельства способствуют усиленному включению глюкозы в биосинтез АК. Усиленное образование АК и НАДФН2 способствуют стабилизации ОВП и предотвращают окисление ГSH. Чему способствует также и высокий уровень таурина, что было показано в наших исследованиях при воздействии на крыс ксенобиотиков – ПАУ, кровопотере, заражении МБТ, радиации, эмоционального стресса. Таурин, ГSH, полиамины накапливаются в кроветворной, лимфоидной тканях обеспечивающих пластическую реконструкцию поврежденных тканей и структур и обеспечивает высокую устойчивость к повреждающим факторам [238].
Однако при повреждающих воздействиях окружающей среды особенно мутагенных и канцерогенных и высоком уровене АФК, радикалов промежуточного метаболизма ГК в тканях (других агрессивных продуктов метаболизирующихся циклических БАВ, а также поступающих извне ПАУ или других мутагенных и канцерогенных ксенобиотиков), несмотря на способность крыс противостоять токсическим и острым стрессовым воздействиям, проявляется отчетливое мутагенное воздействие на генетический аппарат крысы, несмотря на высокий уровень метаболитов-адаптогенов в тканях [358, 359, 368, 298].
Другой тип метаболических реакций на стресс-воздействие наблюдается у морской свинки, которая считается ГК (–) отрицательной. То есть ее рецепторный аппарат (к воздействию ГК) в виде белков-рецепторов (I, II, III) во многих тканях либо мало выражен, либо вообще отсутствует при этом количество цитохрома Р-450 в печени морской свинки в 2-3 раза меньше чем в печени у крыс и соответственно меньше мощность ОСФ. Поэтому метаболические превращения ГК идут в несколько раз менее интенсивно, чем у крысы, и соответственно, других эу- и ксенобиотиков. Поэтому образование АФК и активных радикалов метаболизируемых веществ в печени и других тканях, имеющих системы детоксикации ОСФ, в несколько раз меньше, чем у крысы [227].
Возможно именно эти обстоятельства определили другой спектр метаболитов-адаптогенов, которые образуются в печени морской свинки при стресс-воздействиях (вследствие низкой активности декарбоксилаз, стимулируемых АФК) – это глутамат, пролин, орнитин, глицин, цистеин и образуемый из этих аминокислот глутатион, который в условиях стресс-воздействиях интенсивно окисляется до ГSSГ и никуда из печени не транспортируется [238].
Кроме окисления глутатион интенсивно коньюгируется с метаболитами обезвреживания ГК, КХА, ПАУ и других эу- и ксенобиотиков с удалением этих веществ из клеток и тканей организма морской свинки.
У крыс мощная грубоволокнистая СТ, принимающая активное участие в процессах детоксикации – адсорбирует на своих волокнах токсичные вещества. Клетки СТ крыс способны производить большое количество УДФГК и ФАФС-веществ, коньюгирующих токсичные вещества и выводящие их из организма. Но не все токсичные вещества подвергаются такой коньюгации. Так, производные обмена ГК и ПАУ подвергаются гораздо более эффективной коньюгации и выведенной из организма с глутатионом, как это имеет место у морских свинок. У последних СТ нежноволокнистая синтезировать АК не способна, но при стресс-воздействиях она способна мобилизировать большие количества глутаминовой кислоты, глицина, пролина, необходимых для адаптивных перестроек в печени, прежде всего для биосинтеза глутатиона [227, 86, 238].
Неспособность синтезировать АК приводит к тому, что при стресс-воздействии усиливается обмен углеводов не в сторону образования АК и УДФГК, а в сторону образования рибозы и включения ее в ГМФШ.
Смысл этих превращений состоит в том, что образующихся в ГМФШ НАДФН2 отдает свои восстановленности НАДН2, которые окисляются в ЦТК и дыхательной цепи. Таким путем во всех тканях организма стимулируется пентозный цикл, необходимый для поддержания в восстановленном состоянии глутатиона [170].
Такой ход адаптивных метаболических перестроек без усугубления ОВП и образования АТФ возможен только при интенсивном функционировании ЦТК и дыхательной цепи, то есть усиления метаболических окислительных процессов углеводов по пути превращений в цикле Эмбдена-Меергофа-Кребса с интенсификацией работы ДЦ.
В тоже время у крыс в условиях стресса окисление углеводов по этому пути блокируется и направляется в активированный ГМФШ, обеспечивающий интенсификацию процессов окисления эу- и ксенобиотиков в ОСФ и образование повышенных количеств АФК для усиления свободнорадикального окисления в тканях свободных жирных кислот (СЖК), повышенного биосинтеза АК и метаболитов, используемых в биосинтезе соединительной ткани.
Следовательно, у морских свинок усиленное образование глутатиона, его использование в антиоксидантных и детоксикационных механизмах во всех тканях организма наряду с усилением метаболизирования углеводов по пути Э-М-К и интенсификацией работы ДЦ – являются мощными антимутагенными факторами, что, видимо, и делает морскую свинку устойчивой к мутагенезу и канцерогенезу, а также к хроническому эмоциональному стрессу. По своим механизмам ЭС протекает с усиленным образованием КХА и в гораздо меньшей степени (хотя и постоянно) ГК (т.е. ПАУ) и в процессе их метаболизирования образуются повышенные количества АФК в тканях и токсичные, повреждающие генетический аппарат метаболиты, что напоминает и мута- и канцерогенные воздействия ПАУ [298, 244, 212, 368, 369, 358, 359].
Нам неизвестно по каким причинам морская свинка (в отличие от других видов млекопитающих) избрала этот путь адаптивных перестроек, очевидно, во многом это связано с особенностями обитания ее на островах недалеко от Южной Америки, но у приматов и человека, также не способных синтезировать АК и ориентирующихся в детоксикационных механизмах на повышенное образование глутатиона (его окисление и включение в процессы коньюгации) такая стратегия адаптивных перестроек, очевидно, связана с повышением устойчивости к ЭС, мута- и канцерогенезу. Именно эти обстоятельства, видимо, и определяют тот факт, что на протяжении десятков тысяч лет человек генетически несущественно меняется в отличии, например, от грызунов, собак, копытных и других животных [368, 369, 358, 359].
Имеются сведения, из которых следует, что интенсивность образования таурина в печени снижается в следующем ряду животных: крыса – собака – корова – лошадь – кошка – морская свинка – человек [541], то есть обратно пропорциональна устойчивости к ЭС мута- и канцерогенным воздействиям.
Как выше отмечалось, ткани морской свинки (печень, легкое, селезенка, мозг) содержат железа и серы в 2-3 раза больше, чем соответствующие ткани крысы. Повышенное содержание железа и серы обнаружено в мозге, печени приматов и человека. Железо и сера сосредоточены в основном в митохондриях аэробных тканей [361] в виде железа – серных белков и цитохромов ДЦ. Высокое их количество говорит об ориентации метаболизма на путь Э-М-К с высокой активностью ДЦ и в условиях стресс-воздействий об активизации, а не угнетения этих систем.
Наиболее аэрируемой тканью организма является мозг, содержащий наибольшее количество митохондрий и железосерных комплексов по сравнению с другими тканями организма [235,22]. Считается, что обмен нервной и мозговой ткани в принципе одинаков у всех животных [158]. Однако в условиях стресс-воздействий у различных видов животных аэрация мозга различна, как различны и субстраты для энергообразования [244].
Так, мозг крысы в условиях стресс-воздействия вполне может окислять СЖК с образованием большого количества АТФ, и достаточно устойчив к глубокой гипоксии. В тоже время мозг человека, приматов и морской свинки в условиях стресс-воздействия облигатно зависим от поступления углеводов и достаточной степени аэрации, особенно при ЭС.
При этом необходимо указать, что внутри каждого вида имеются особи существенно отличающиеся друг от друга по чувствительности к различным стресс-воздействиям.
Из имеющихся литературных данных [244] известно, что животные, также как и человек могут относится к разным защитно-адаптивным типам. Так, выделяют высоко устойчивых особей (как между видами так и внутри каждого вида) к острым и интенсивным стресс-воздействиям, которые гораздо менее устойчивы к воздействиям менее интенсивным, но длительным. Это так называемые «спринтеры». Другой крайний тип – это особи устойчивые к длительным и мало интенсивным воздействиям (особенно к эмоциональному стрессу), но менее устойчивые к острым и интенсивным стресс воздействиям (особенно физическим повреждающим факторам – травма, температура, токсические воздействия и т.д) — “стайеры”. Между этими типами существуют различные промежуточные типы [138, 294, 268, 386, 322, 323].
Нам удалось установить [238], что для «спринтеров» более характерен N-тип защитно-адаптивных механизмов (печеночный соматический метаболический тип с преобладанием в адаптивных механизмах метаболитов богатых азотом), ориентирующийся в основном на перестройку метаболических процессов в печени с выделением в кровь таких веществ как таурин, аргинин, полиамины, мочевина, то есть веществ, содержащих в повышенных количествах азот. Из метаболитов, содержащих серу при таком защитно-адаптивном типе, преобладает образование таурина, в котором сера окислена до SO3.
Для «стайеров» более характерно наличие S-типа с большим значением защитных механизмов перераспределения аминокислот из соединительной и нервной ткани. Здесь преобладают такие метаболиты как глутаминовая кислота, пролин, глицин, цистеин и глутатион, то есть вещества, содержащие в повышенном количестве серу, в которых она находится в восстановленном состоянии: SH.
Выше уже отмечалось, что стресс-воздействия влияют на защитно-адаптивные системы организма, стимулируя их. Соответственно повышается устойчивость к повреждающим воздействиям (эустресс). И напротив, когда происходит истощение защитно-адаптивных систем, развивается патологическое состояние (дистресс). Частые, превышающие адаптивные ресурсы, повреждающие воздействия могут привести к гибели организма.
Организмы с преобладанием N-типа (“крысы”) более устойчивы к острым и интенсивным стресс-воздействиям (перепады температур, физическая нагрузка, возбудители инфекционных заболеваний и т.д.). Это связано с тем, что вещества-адаптогены, образующиеся в повышенном количестве при таком защитно-адаптивном типе, обладают мембраностабилизирующим эффектом и рядом других особенностей, необходимых при грубых и интенсивных стресс-воздействиях. То есть этот защитно-адаптивный тип направлен на “противоудар” — на сопротивление.
Организмы с преобладанием S-типа защитно-адаптивных систем малоустойчивы к интенсивным и острым, но гораздо более устойчивы к малоинтенсивным и длительным стресс-воздействиям. Малоинтенсивный, но хронический стресс требует соответствующей ему стратегии защиты. Важнейшее значение имеет то обстоятельство, что такие стресс-воздействия сопряжены с постоянным повышением в тканях активных форм кислорода и свободных радикалов (хотя и в гораздо меньших количествах, чем при острых и интенсивных стресс воздействиях), которые оказывают повреждающее воздействие на мембраны клеток, особенно митохондрий, ответственных за образование АТФ и обеспечение организма энергией. Для защиты клеток в такой ситуации необходимы вещества, образующиеся в большом количестве в мозге и нервной ткани. Из этих веществ-адаптогенов образуется глутатион, который и является главным фактором защиты при хроническом стресс-воздействии. Экспериментально доказано, что наибольшее отрицательное влияние оказывает хронический эмоциональный стресс, при котором повреждаются как митохондрии, так и ядерный аппарат клетки [244]. Это приводит к повышению количества мутаций и канцерогенным воздействиям, что сопровождается развитием различных патологических состояний, вплоть до возникновения злокачественных опухолей [359]. Главным защитным механизмом в данной ситуации является глутатион. На этой основе формируется другой тип защитно-адаптивных систем — тип “избегания”, когда защитно-адаптивные системы направлены не на сопротивление повреждающему воздействию, а на уход от него. Главенствующее значение в такой стратегии имеет развитый мозг и другие отделы нервной системы.
Таким образом, развитие высокоорганизованной нервной системы является средством приспособления организма к хроническим стресс-воздействиям, прежде всего к эмоциональному стрессу, поскольку именно в нервной системе и образуются вещества (глутаминовая кислота, цистеин, глицин), необходимые для образования глутатиона. Эмоциональный стресс в силу этой причины является мощным фактором эволюционного развития, так как является фактором стимуляции цефализации, существенно ускорившей эволюцию. Так, в определенной мере, эволюционное восхождение в плане совершенствования адаптивно-защитных механизмов среди млекопитающих можно проследить от грызунов (крысы, мыши) и до приматов.
Известно, что крыса является своеобразным высоко адаптивным биологическим реликтом (Ж. Сент – Илер 1968), хорошо переносящим острые и интенсивные повреждающие воздействия (температура, токсические вещества, радиация и т.д.). Важнейшее значение в защитных механизмах к повреждающим воздействиям у крысы имеет печень, где в большом количестве образуются адаптогены (аргинин, полиамины, мочевина и ряд других), которые накапливаются в важнейших защитно-адаптивных системах организма (кроветворная, иммунная, детоксикационная и др), но особенно в больших количествах в тканях у крыс обнаруживается таурин — 50% и более от всех аминокислот. Промежуточное положение занимают другие млекопитающие. Так, например, если у крыс содержание таурина максимальное и максимальная устойчивость к острым и интенсивным стресс-воздействиям, то, для сравнения, у собак содержание таурина гораздо меньше, и меньшая устойчивость к аналогичным стресс-воздействиям. Еще меньшая концентрация таурина у кошек. Одно из последних мест в этом ряду (по количеству таурина в тканях) занимают приматы и человек. Но у человека имеются два полюса защитно-адаптивных систем-N-тип и S-тип, что делает его гораздо более адаптивным по сравнению со всеми живыми существами по отношению к воздействиям факторов окружающей среды.
Таким образом, завершает эту пирамиду человек, имеющий максимальные защитно-адаптивные возможности. Эти возможности состоят из двух полюсов- “печеночного” (N-тип) и мощного “мозгового” (S-тип), что позволяет иметь среди человеческой популяции широкий спектр различных вариантов защитно-адаптивных механизмов. В плане экспериментальных исследований защитно-адаптивных типов большую ценность имеют опыты, проведенные нами на крысах и морских свинках. У этих животных защитно-адаптивные типы являются как бы крайними вариантами, присущими для человека.
Про крыс мы уже упоминали – это крайний печеночный N-тип. Другим крайним типом является морская свинка. По своим обменным процессам, в силу некоторых обстоятельств, похожа на приматов и человека, что могло быть обусловлено особенностями ее жизни на островах, в определенных условиях. Морская свинка имеет ярко выраженный S-тип (перераспределительно-мозговой), но мозг у нее очень маленький, а “печеночный” N-тип выражен очень слабо. Для морской свинки характерна повышенная чувствительность к различным острым и интенсивным стресс-воздействиям (радиации, перепадам температур, интоксикациям, заражению многими возбудителями инфекционных заболеваний и т.д.). Но она имеет высокую устойчивость к факторам, повреждающим ее генетический аппарат (хронические интоксикации, мутагенные и канцерогенные воздействия).
Важнейшими факторами, определяющими устойчивость, как морской свинки, так и человека к хроническим малоинтенсивным стресс-воздействиям, эмоциональному стрессу, мутагенным и канцерогенным воздействиям являются образующиеся в их организме, в большом количестве, вещества, защищающие генетический аппарат от мутагенных (вызывающих мутации) и канцерогенных (вызывающих опухоли) веществ. К таким защитным метаболитам относятся глутатион, цистеин, и ряд других, серосодержащих аминокислот и их производных. Такие метаболиты или их предшественники, в большом количестве, образуются в мозге и нервной системе. А также они содержатся в соединительной ткани, из которой при стрессовых воздействиях могут перераспределяться в защитно-адаптивные системы организма (перераспределительный тип).
Чем более развит вид в плане совершенствования его ЦНС и мозга, тем более различна реакция его представителей на то или иное стресс-воздействие. Так, мы уже отмечали, что исследователи выделяют «спринтеров» и «стайеров» по устойчивости к острому и интенсивному и малоинтенсивному, но хроническому стрессу [138, 383, 365].
Однако необходимо констатировать, что в принципе все млекопитающие, находящиеся в диких условиях природы, с каждым годом все более и более теряют свою адаптивность и исчезают как виды.
Это касается также и многих других представителей флоры и фауны. Все это происходит вследствие колоссальной антропогенной нагрузки и чрезвычайно высокой адаптивности как вида человека. Но по мере нарастания техногенной нагрузки на биосферу и загрязнения окружающей среды выбросами различных производств, транспорта, отходами производства и быта людей они и сами начинают терять адаптивность. Главный показатель падения адаптивности – резкое падение рождаемости, особенно технически высокоразвитых народов.
Здесь необходимо остановиться на том, что человек, имеющий наиболее развитую ЦНС и мозг среди всех других представителей живой природы, внутри своего вида имеет максимальные различия по отношению к воздействию различных стресс-факторов [244, 138].
Так, из многочисленных научных исследований да из наблюдений в быту хорошо известно, что отдельные люди хорошо переносят острые и интенсивные стресс-воздействия, то есть являются «спринтерами», а другие – малоинтенсивные, но длительные — хронические – «стайеры». Так, среди «спринтеров» можно отметить людей, хорошо переносящих физическую нагрузку, они, как правило, хорошо концентрированы, имеют мощные тормозные процессы на уровне ЦНС – хорошо засыпают, уравновешены, внимательны [138, 383, 365].
Такие люди чаще устойчивы к интенсивной физической нагрузке, холоду, инфекционным заболеваниям, воздействию токсичных веществ. Более того, они склонны к употреблению алкоголя и курению. Но они чаще всего не склонны к интенсивной умственной деятельности, плохо переносят хронический эмоциональный стресс – следствием чего является алкоголизм, психические расстройства.
Другой крайний тип «стайеры» – это люди чаще умственного труда, плохо переносящие острые и интенсивные физические стресс-нагрузки, имеют слабые тормозные процессы на уровне ЦНС, не склонны к употреблению алкоголя и курению. Однако такой тип защитно-адаптивных механизмов устойчив к хроническому эмоциональному стрессу, высоким умственно-коммуникативным нагрузкам [244, 294, 204, 320].
Согласно данным токсикологических и патофизиологических исследований повышенную устойчивость к различным интоксикациям и повреждениям имеют люди с мощной системой оксигеназы смешанных функций ОСФ и повышенным количеством Р-450 в печени, функция которых обеспечивается мощным ГМФШ в печени [86].
Таким образом, люди, являющиеся в большей степени «спринтерами», ориентируются в своих защитно-адаптивных механизмах на доминирующую роли метаболизма печени, тогда как у «стайеров» в большей мере доминирует метаболизм мозга и в целом надо констатировать, что у человека в отличии от животных имеются как бы два полюса образования метаболитов-адаптагенов – печень и мозг. Это определяет широкий спектр защитно-адаптивных типов в человеческой популяции и делает ее гораздо более адаптивной по сравнению с популяциями и тем более отдельными особями всех других видов.
Воздействие на организм вирулентной микрофлоры при развитии и прогрессировании инфекционного процесса (например, мышиный тиф, туберкулез, трипаносомоз и др.) снижает уровень цитохрома Р-450 в печени и наоборот относительно легко перенесенная инфекция и выздоровление повышают уровень и соответственного детоксикационные ресурсы организма. В печени образуются белки системы комплемента, существенно повышающие антимикробные свойства иммунной системы [248, 378].
Точно также и воздействие мута-канцерогенных ксенобиотиков типа ПАУ в небольших субтоксичных для данного организма дозах повышает иммунные и детоксикационные ресурсы [149]. То есть людей такого типа условно можно назвать «печеночники».
И, напротив, в условиях хронического эмоционального стресса главенствующие значения в ЗАМ имеют не детоксикационно-метаболические ресурсы печени, которые включают в себя системы ОСФ – цитохром – Р-450 с интенсивным биосинтезом таких веществ как таурин и стабилизаторы мембран — полиамины, а такие адаптогены как глутамат, ГАМК, ГОМК, карнозин, гамма–глутамил пептиды (например, Г-SH), аргинин, мочевина, защищающие, например, миокард от повреждающего воздействия [202]. Все эти вещества образуются в головном мозге в повышенных количествах и в условиях стресс-воздействия могут защищать как саму ткань мозга, так и перераспределяться в другие органы (для чего есть специальные транспортные системы). Оказывают защитный эффект, например, в сердечно-сосудистой системе, миокарде [264].
То есть, люди с преобладанием такого защитно-адаптивного типа механизмов являются «мозговиками».
Однако деление это условно, так как мозг и печень — 2 наиболее крупных паренхиматозных органа, которые составляют примерно по 2% от массы тела человека, дополняют и координируют функцию друг друга в условиях адаптивного ответа на стресс-воздействие. Так, печень обеспечивает мозг серосодержащими метаболитами-аминокислотами, для которых в ГЭБ существуют особые транспортные системы. Таурин и цистеин активно поглощаются мозговой тканью. Таурин обладает мощным общим тормозным действием, координирующим многие медиаторно-регуляторные эффекты в коре и подкорковых структурах. Мозговая ткань содержит большие количества серосодержащих аминокислот – таурина, цистеина, цистатионина, изетеиновой кислоты [264, 225].
В свою очередь в головном мозге образуется большое количество – азотсодержащих метаболитов – адаптогенов широкого спектра – глутамата и его производных ГАМК, глутамин, мочевина, N-ацетилглутамат, глутамин транспортирующихся в печень и стимулирующих здесь адаптивные процессы, например, биосинтез мочевины стимулируемый N-ацетилглутаматом, биосинтез нуклеиновых кислот и полиаминов, стимулируемых N-ацетиласпартамом и глутамином, поступающих из мозга и т.д. [158, 176, 264, 386].
Однако, несомненно, и то, что в адаптивных процессах в зависимости от условий среды, в ответ на стресс-воздействие может преобладать либо мозг, либо печень и соответственно люди с таким преобладанием могут условно обозначаться – «мозговики» или «печеночники».
В проведенных нами исследованиях спектра аминокислот-адаптогенов у 5% людей он напоминает таковой у морской свинки. У 42% он напоминает таковой у крысы. 53% различные промежуточные типы.
В первом случае это были чаще всего люди умственного труда, с относительно слабым здоровьем. Если они заражались туберкулезом, то он протекал у них тяжело, а лечение противотуберкулезными препаратами часто давало побочные эффекты в виде гепатитов, поражений нервной системы, слухового аппарата и т.д.
Во втором случае – это чаще всего люди физического труда, не склонные к напряженной умственной и творческой деятельности. У людей второго типа туберкулез развивался либо после длительного пребывания в заключении или воздействия других сильных и длительно действующих стрессовых факторов.
Соотношение аминокислот адаптогенов и некоторых их производных, которые определяются в крови здоровых и больных различными заболеваниями людей, дает определенные коэффициенты. По этим коэффициентам можно рассчитать принадлежность индивидуума к тому или иному адаптивному типу и метаболический адаптивный ресурс организма. Учет полученных результатов позволил назначить и провести целенаправленную коррекцию метаболических и адаптивных нарушений и существенно повысить эффективность лечения, например, тяжелых форм туберкулеза [238].
Полученные результаты позволяют говорить о том, что у людей, спектр аминокислот-адаптогенов которых в определенной степени соответствует таковому у крысы, эти вещества нарабатываются в основном в печени. У людей спектр аминокислот-адаптогенов в крови напоминающих таковой у морской свинки эти метаболиты образуются в основном в мозге, поэтому первых условно можно обозначить как «печеночники», а вторых как «мозговики». Люди 2-го типа – чаще всего люди умственного труда, чувствительные к инфицированию МБТ, люди 1-го типа – чаще всего люди физического труда, заболевают туберкулезом лишь после длительных неблагоприятных условий жизни – длительный стресс, длительные сроки заключения [238].
Весьма интересны сведения о становлении человека как обладателя высокоадаптивного типа защитных механизмов.
Так, известным русским психологом, физиологом, антропологом и историком Поршневым Б.Ф. была выдвинута достаточно аргументированная концепция о том, что промежуточный этап перехода от обезьяны к человеку имел те особенности, что существенно менялись форма и объем мозга. И если у наших предков, еще целиком находящихся в царстве животных, размеры мозга (понгиды) соответствуют примерно современным шимпанзе, то уже следующий этап перехода – троглодиты размеры мозга значительно увеличиваются, доходя до максимальных размеров у палеоантропов и неоантропов, что можно видеть из приведенной таблицы (1.8), взятой из монографии Поршнева Б.Ф. (по Кочетковой В.И.)
Табл.1.8 Размеры мозга по эндокранам ископаемых черепов:
| понгиды | троглодиты | гоминиды | ||||
| В мм | шимпанзе | австралопитек | Археантроп (питекантроп) | Палеантроп (неандерталец) | Неантроп (кроманьонец) | Современный человек |
| Длина мозга | 106 | 125 | 155 | 174 | 186 | 167 |
| Лобная доля | 81 | 92 | 110 | 128 | 135 | 129 |
| Теменная доля | 71 | — | 108 | 118 | 116 | 124 |
Показатели, составленные по эндокранам (вдавлениям) ископаемых черепов имеют самый большой размер у неоантропов (кроманьонцев) – ископаемых гоминид. И он уже достаточно большой у предшествовавших им троглодитов – палеоантропов (неандертальцев), еще собственно к людям (гоминидам) не относящихся. Но, как видим, и у тех и у других меньшие размеры теменной доли, в которой сосредоточены речевые центры. Удивительно мощное адаптивное значение мозга (наработка метаболитов-адаптогенов) его значительное увеличение у неандертальцев и особенно кроманьонцев имело ведущее значение в совершенствовании механизмов адаптации.
ГЛАВА 2 ОСОБЕННОСТИ ОБМЕНА АМИНОКИСЛОТ У СПОРТСМЕНОВ РАЗЛИЧНЫХ СПЕЦИФИКАЦИЙ
2.1 ВВЕДЕНИЕ.
Классификации конституции включают в себя как морфологические, так и физиологические признаки. Кроме того, глубокое исследование человеческого тела позволяет выявить великое множество индивидуальных особенностей анатомического строения. Размеры тела и его частей или органов, их пропорции и соотношения не идут ни в какое сравнение с количеством вариантов кровоснабжения частей тела или органов, что послужило причиной множества анатомических исследований, позволяющих избежать врачебных ошибок, часто трагических. Врачебные классификации Черноруцкого, Шелдона, Кнрада и др. с давних пор помогали врачам по внешним признакам, сочетаниям размеров тела, его компонентного состава составить представление о ходе болезни, особенностях течения патологических процессов и о возможностях излечения. Таким образом давно уже установлено, что внешние данные тела, особенности его строения, особенности кожных покровов имеют связь с особенностями реагирования как на заболевание, так и на ход излечения от этого заболевания.
Классические исследования Кречмера, Шелдона и других прямо показывают, что разные аспекты человеческой сущности – морфологические, физиологические биохимические и психологические связаны между собой. Кречмер установил связь внешних признаков и склонность к определенным психическим отклонениям. Многоплановые исследования Шелдона не только установили связь между преобладанием компонентов тела и психологическими характеристиками человека, но и указал на единство происхождения морфологических и психологических характеристик в преобладании того или иного зародышевого листка. Эндоморфия находит свое выражение в висцеротонии, мезоморфия – в соматотониии эктоморфия – в церебротонии.
Последующие исследования (Мерлин) посвященные исследованию целостности человеческой сущности показали в качестве основы единства индивидуальный стиль деятельности, что хорошо согласуется с теорией функциональных систем (Анохин), но ничего не говорит о материальном субстрате, формирующем это единство.
Представленные в первой главе материалы говорят о том, что таким субстратом могут быть обменные процессы, нашедшие свое выражение в аминокислотном составе плазмы крови.
2.2 ОБЗОР ЛИТЕРАТУРЫ ПО АМИНОКИСЛОТАМ У СПОРТСМЕНОВ
Современный спорт высоких достижений требует грамотной и эффективной фармакологической поддержки [207, 133, 134, 160, 161, 162, 163, 187, 292, 299, 234, 103, 104, 302].
Для чего используется большое количество различных препаратов, повышающих адаптивные возможности организма к физическим нагрузкам в условиях других стрессовых воздействий на организм при занятиях спортом и соревнованиях [207, 164, 165, 166].
В большинстве исследований по изменению метаболических и функциональных возможностей органов и тканей спортсмена установлено, что интенсивная физическая нагрузка сопряжена с усилением свободнорадикальных процессов в мышцах и других тканях организма [29, 30, 284, 403, 405, 128, 217, 303, 315, 402, 484, 499, 515, 533, 534, 577, 588]
Интенсивная мышечная деятельность приводит к перераспределению крови и в значительной степени перераспределению важнейших метаболитов, характерных для обмена веществ при физической работе, необходимых для жизнедеятельности организма. Происходит перестройка нейроэндокринной, кроветворной, дыхательной, выделительной систем оргавнизма. Однако все эти изменения имеют свои особености как в различных видах спорта так же как зависят от особенностей организма спортсмена в реагировании на физический и эмоциональный стресс и другие виды стресса, воздействующие на спортсмена. Исследованию морфологических, физиологических, психологическиъх особенностей спортсменов и вообще воздействию физической и психологической нагрузки на организм человека и животных посвящено большое количество исследований. Поэтому имееется достаточно полная картина перестроек систем организма при различных спортивных нагрузках и в различнм возрасте [14, 15, 17, 18, 48, 60, 62, 63, 64, 80, 107, 108, 109, 112, 114, 154, 155, 156, 169, 171, 173, 174, 193, 194, 195, 196, 197, 198, 260, 242, 261, 283, 288, 306, 307, 308, 309, 310, 311, 313, 314, 319, 320, 321, 322, 323, 324, 325, 326, 327, 328, 339, 338, 376, 378, 352, 353, 354, 355, 356, 366, 386, 387, 388, 389, 390, 391, 395, 53, 57, 495].
Работа мышц, так же, как и работа нейронов мозга, сопряжены с генераций большого количества свободных радикалов кислорода и азота [41, 517, 518, 569, 599, 585, 599, 537, 538], аммиака, глутаминовой кислоты [503, 576], подлежащих интенсивной детоксикации, так как в повышенных количествах и аммиак и глутамат токсичны для нейронов [22, 191, 215]. Для этого существуют мощные системы защиты от свободных радикалов и токсичных метаболитов, образующихся при физической работе [517, 518, 519, 520, 522].
Чем выше уровень тренированности спортсмена, тем более мощные системы антиоксидантной защиты и детоксикации токсичных веществ, образующихся в ходе интенсивно протекающего метаболизма при повышенных физических нагрузках [41, 174, 527, 528, 529, 525, 531, 532].
Помимо чисто физическго напряжения спортсмен испытывает психологический стресс различной степени интенсивности. Издесь уже важны особенности темперамента человека, его психологические особенности.
Темперамент издавна рассматривался с точки зрения выявления наилучших предпосылок для той или иной деятельности. Давно установлены темпераменты, наилучшие для многих видов спорта. При этом люди, имеющие темперамент, известный, как совершенно неподходящий для данного вида спорта порой занимаются этим видом не только успешно, но и добиваются высоких результатов [2]. Тем не менее, исследование темперамента обычно происходит с целью отнести спортсмена к той или иной группе, и на основании принадлежности к группе холериков, меланхоликов, сангвиников или флегматиков, либо определения базовых свойств нервной системы строится диагностика годности к виду спорта, прогноз на результаты деятельности и особенности педагогической работы с данным спортсменом.
Влияние спорта на организм многообразно, и психологические изменения входят в список последствий спортивной деятельности. Какие психологические изменения происходят под влиянием спорта вообще, и конкретного вида спорта в частности, в целом известно [1, 4]. В то же время остается неизвестным, как влияют занятия подходящим по темпераменту видом спорта, и занятия видом спорта неподходящим для данного темперамента. Выделить то или иное влияние на организм спортсмена довольно сложно. Известно, что спортсмены, достигшие определенного уровня мастерства, отличаются от неспортсменов не только физической силой, но и морально-волевыми качествами, способностью к преодолению не только собственной слабости, но и к преодолению воли соперника. Упорство в достижении цели, характерное для большинства видов спорта должно сочетаться с противоречащими качествами — быстротой реакции, и быстрого поиска новых решений в мгновенно возникающих и исчезающих ситуациях [7, 200, 260, 287, 294, 302, 326, 327, 331, 335, 341, 343, 344, 346, 376].
Предыдущая глава была посвящена тому, что особенности обмена аминокислот, их распределение и количество (а так же ряда их производных таких как глутатион, полиамины, ГАМК, анзерин, карнозин, адреналовые и тиреоидные гормоны, нейропептиды, креатинфосфат и другие биологически активные метаболиты) в тканях организма у животных и человека во многом определяют их адаптивный тип и соответствующую реактивность на стресовые воздействия. Ведущее значение в обеспечении организма перечисленными адаптогенами и в реализации их биологического эффекта имеют мозг и печень. При преобладании в адаптивных механизмах мозга (здесь аминокислот и ряда их биологически активных производных-адаптогенов на порядок, а то и на два больше, чем во всех других органах и тканях) определяет преимущественно «МОЗГОВОЙ» адаптивный тип, который видимо будет лучше рализовываться в определеных видах спорта. Имеются сведкения, что перечисленные метаболиты влияют на особенности обменных процессов и адаптивных перестроек при спортивных нагрузках [594, 593].
Другим важным органом обеспечивыющим организм адаптогенами является печень. Здесь синтезируются такие важные для устойчивости спортсмена к физическим и психологгическим нагрузкам метаболиты как таурин, глутамат, аланин [593, 595].
В печени сосредаточено основное количестьво метионина в организме, и она во многом определяет обмен серы в организме, то есть обмен и количество в тканях аминокислот-адаптогенов, содержащих серу. В печени происходит детоксикация аммиака, молочной кислоты, обезвреживание образующихся в мышцах и мозге при их интенсивной работе, а также обезвреживание избытка гормонов, образовавшихся при физическом и эмоциональном стрессе перживаемом спортсменом во время тренировок и соревнований [581]. Помимо того катаболические процессы стрессового воздействия сопряжены с увеличением в тканях организма не только метаболитов необходимых для дальнейших структурных перестроек (аминокислоты, нуклеотиды, липиды и фосфолипиды и т.д., но и различных токсичных продуктов деградации клеток подверженных «реконструкивному протеолизу». За детоксикацию и выведение из организма таких продуктов так же ответственна печень [155, 166, 168].
Мозг, печень, мышцы, соединительная ткань составляют как бы единую адаптивную систему по производству, депонированию, метаболизированию и использованию в качестве адаптогенов дикарбоновых кислот. Прежде всего это касается таких метаболитов как глутамат и глутамин.Но в этой системе может преобладать либо то, либо другое. Поэтому индивидуумы, обладающие более мощной печенью, имеют скорее «ПЕЧЕНОЧНЫЙ» тип адаптации и для них вероятно более подходят другие виды спорта, чем для «МОЗГОВИКОВ». Вероятно, можно выделить и «МЫШЕЧНЫЙ» адаптивный тип. Но соответствующих исследований ранее не проводилось. В то же время такие исследования могли бы иметь большую ценность так как позволяли более целенаправленно производить отбор в спорт, более рационально выстраивать тренировочный режим и при этом целенаправленноо и с гораздо большим эффектом использовать парепараты-адаптогены, прежде всего аминокислоты и их производные. Все изложенное позволяет сделать следующие предположения.
Метаболизм мозга связан с метаболзмом соединительной ткани, ибо глия, во многом обеспечивающая метаболические потребностим нейрона, во многом состоит из соединительной ткани и нарабатывает вещества необходимые нейронам (глутамат, глутамин, глицин), а также депонирует метаболиты, используемые нейронами при усилении их деятельности [22].
Глутаминовая кислота, пролин, таурин, таким образом, могут быть метаболитами связывающими работу мозга, работу ОДА, через метаболизм соединительной ткани. Переразвитие мышц у борца или хоккеиста приводит к переразвитию упругого мышечного компонента – соединительнотканного каркаса. Отсюда реконструктивный протеолиз не может быть чрезмерным, как у легкоатлетов, и на определенном уровне мастерства суммарный аминокислот снижается.
Большая мышечная масса образует много аммиака и молочной кислоты и глутамина. Должна быть мощная печень, чтобы стабилизировать гомеостаз.
Большая мышечная масса, не скоординированная с работой печени и мозга, при работе становится токсичной для организма – аммиак, молочная и глутаминовая кислота вместе создают токсический гомеостаз. Видимо с этим связано большое количество всяких патолгических процессов у спортсменов, имеющих большое количество мышечного компонента. Что усугубляется употреблением большого количества всяких анаболиков (594). Длительные занятия спортом приводят к изменениям в организме, как функциональным, так и анатомическим. Формирование структурного следа адаптации к физическим нагрузкам очевидно для внешнего наблюдателя и выражено в словах «спортивная фигура». Необходимость изучения морфологических особенностей тела человека в связи с занятиями спортом обусловило появление спортивной морфологии.
Исходя из имеющихся данных для коррекции возможных нарушений перечисленных адаптивных систем при повышении спортивных результатов чаще всего предлогаются антистрессовые вещества неспецифического характера воздействия (женьшень, элеутерококк, левзея и тд. Кроме того, часто используются фармакологические препараты, одновременно обладающие антистрессовым, антигипоксантным и антиоксидантным эффектом (янтарная кислота, глутаминовая кислота, глицин ит.д. [562].
Среди этих препаратов как в спорте, так и в фитнесе наиболее широко используются некоторые аминокислоты- естественные адаптогены, выполняющие в организме различные адаптивно-регуляторные и защитные функции. Большое количество исследований посвящено применению в спорте различных аминокислот. Показаны положительные результаты их применения на повышение устойчивости к стрессу (аспартат, глутамат, глицин, таурин), выносливости (глутамат, цистеин, глицин), силовых (аргинин, глицин, метионин), скоростных качеств спортсмена, показан их анаболический эффект на прирост мышечной массы (разветвленных аминокислот и их комплекса в виде ВСАА, глутамина), повышение иммунных возможностей спортсмена (лизин, аргинин), усиление антиоксидантных ресурсов организма (цистеин, глутамат, глицин) и состоящего из этих аминокислот трипептида глутатиона Подобраны дозы аминокислот рекомендуемых при тех или иных видах спорта [167, 216, 217, 93, 94, 95, 51, 69, 70, 109, 266, 479, 484, 491].
Имеются работы [77, 78, 79, 167, 216, 217], в которых достаточно подробно исследовано изменение количества аминокислот у спортсменов-пловцов. Из этих работ следует, что наибольшее значение в адаптивно- метаболических перестройках у этих спортсменов имеют серосодержащие аминокислоты (таурин, цистенин, цистеиновая кислота, метионин и их производное глутатион). Авторы рекомендуют применять в диете повышенное количество этих метаболитов и витаминов, участвующих в их метаболизме.
Имеется ряд иностранных исследований, посвященных роли свободных аминокислот в метаболических и функциональных перестройках у спортсменов различных спецификаций, отмечается положительное воздействие ВСАА, аргинина, лизина, комплексных препаратов на основе аминокислот, положительно влияющих на тренировочный процесс и спортивеные достижения [588, 402, 533, 534, 536, 577, 424, 425, 421]
Имеющиеся работы чаще всего показывают положительное воздействие аминокислот на какую-нибудь одну из сторон гомеостаза — анаболический эффект, антигипоксантное воздействие, антиоксидантный эффект или на гомеостаз в целом — антистрессорный эффект, улучшения функции сердечно-сосудистой, дыхательной системы и т.д.. Но исследований, касающихся интегративной роли не отдельной аминокислоты, а аминокислотного состава в целом на различные адаптивные системы организма спортсмена, тем более, в различных видах спорта мы не встретили.
В то же время на основании результатов исследований, приведенных и опубликованных нами ранее [237,238,239] и имеющихся данных литературы [22, 23, 96, 132, 133, 134, 181, 215, 243, 260, 306, 311] можно утверждать, что многие аминокислоты и некоторые их производные (ГАМК, ГОМК, биологически активные амины, гормоны и ряд других) обладают комплексным и интегративным воздействием на организм человека и животных. Так в 1 главе мы отмечали, что, например, глутамат, является метаболитом чрезвычайно важным для функции и адаптивных перестроек при стрессе нейроэндокринной, иммунной, мышечной систем, является антигипоксантом, антиоксидантом, анаболиком и обладает еще рядом свойств, которые могут быть важны для спортсмена в условиях повышенных нагрузок. В определенной степени это же касается таких аминокислот, как аспартат, глицин, серосодержащие аминокислоты, циклические аминокислоты. Да по сути дела всех аминокислот вообще и ряда их производных (таких как ГАМК, ГОМК, олигопептиды-глутатион, тафтсин, анзерин и карнозин, и т.д.)
Но в то же время не всегда и не во всех случаях аминокислоты вводмые извне вовышают адаптивные возможности организма, тем более при высоких физических нагрузках и в условиях воздействия различных стрессовых факторов, таких как перемена климата, особенности психологического и физиологического состояния, эмоциональный стресс, и еще множества факторов, действующих в реальной обстановке на спортсмена в условиях тренировок и сорнвнований. Нерациональное применение аминокислот может усугубить отрицательнеое воздействие стрессовых воздействий [237, 238, 239,].
Люди, так же как и животные имеют различныем адаптивные типы [244,193-202,268,237,238,239]. И применение у них одних и тех же аминокислот – адаптогенов может оказывать прямо противоположное воздействие, вплоть до тяжелых отрицательных последствий, что мы и наблюдали в эксперименте на животных, а также при использовании аминокислот у спортсменов. В то же время в литературе мало сведений об отрицательных эффектах используемых аимнокислот. Видимо коммерческий интерес производителей и продавцов аминокислот акцентирует в основном их положительные воздействия.
Всесторонних исследований о роли аминокислот в регуляции важнейших адаптивных систем организма при высоких спортивных нагрузках нами не обнаружено. Что и определило актуальность проведеного исследования.
ЦЕЛЬЮ данного исследоывания как раз и было установить интегративное значение ряда аминокислот и их соотношений на важнейшие адаптивные системы организма спортсмена, а также подтвердить или опровергнуть тот факт, что доминирование определенных аминокислот и их соотношений может определять адаптивный тип человека. Как мы указывали ранее в крайних формах это «МОЗГОВОЙ» и «ПЕЧЕНОЧНЫЙ» типы.
2.3 Материалы и методы
Были взяты 20 борцов и 37 легкоатлетов, 22 хоккеиста Все спортсмены – от перворазрядников до мастера спорта международного класса — ведут одинаковый образ жизни – вся жизнь подчинена тренировочно-соревновательным циклам. Подготовка к соревнованиям, так называемые сборы, проходит в течение 2-3 месяцев, после чего спортсмены участвуют в соревнованиях. После непродолжительного отдыха цикл повторяется. Во время сборов спортсмены проводят ежедневно 2-3 тренировки различной длительности. Таким образом, они заняты преимущественно тяжелым физическим трудом, направленным на совершенствование собственного тела для выполнения определенной, характерной для вида спорта деятельности, чьим содержанием является достижение максимального результата. Факт наличия спортивного звания (1 разряд, кандидат в мастера спорта (КМС), мастера спорта (МС) или мастера спорта международного класса (МСМК) является свидетельством того, что максимальный результат в пределах вида спорта в рассматриваемой группе достигнут примерно одинаковый.
Борцы и легкоатлеты были выбраны как виды спорта абсолютно различные по содержанию деятельности. Представители этих видов спорта выступают в соревнованиях как отдельные личности, противостоящие соперникам различным способом. Борцы непосредственно вступают в силовое противоборство с противником. Каждый борец имеет в своем арсенале набор навыков-приемов, используя которые он стремится повергнуть противника. Противник, естественно, имеет аналогичный арсенал навыков и такое же стремление. Соответственно, на каждое действие противник постоянно находит и использует навык-противодействие, и поиск такого противодействия не занимает много времени – опоздание означает поражение. Приемы могут быть отработаны до автоматизма или придуманы ex tempore в ходе схватки. Каждый прием, отработанный до автоматизма, можно рассматривать как разновидность фиксированной формы поведения (ФФП). В то же время поведение противника постоянно вносит коррективы в деятельность борца, и форма его противодействия может быть эффективной как контрприем, то есть не просто противодействие, но и действие, способствующее победе противника над спортсменом. Противники спортсмену встречаются разные, с разными характерами, каждый со своими излюбленными приемами. Постоянной для спортсмена величиной является его собственное тело и механизмы управления им. Непостоянство же внешней среды – противника предъявляет спортсмену требование сочетания ФФП с предельно возможной гибкостью поведения в соответствии с индивидуальными особенностями противника.
Борцу тактику борьбы диктует собственное тело – и тело противника. У борца есть две тенденции – использовать ФФП и быть предельно гибким поведении в поединке. Предварительное продумывание тактики и использование заготовленного тактического построения борьбы сталкивается с аналогичным построением и, при любой эффективности не исключает поиска решения ex tempore, самостоятельно в критической ситуации.
Легкоатлеты в нашем исследовании – бегуны. Мы заведомо исключили прыгунов и метателей, в силу их специфичности и малодоступности, а бегунов — мастеров спорта найти легче, мы их и нашли. Легкоатлеты уровня мастера спорта ведут образ жизни неотличимый от борцов – такие же тренировочные циклы, сборы, соревнования. Но в поведении легкоатлеты разительно отличаются от борцов. Обращает на себя внимание их привязанность к тренеру. Видимо дело в том, что легкоатлеты в соревнованиях действуют хотя и самостоятельно, но тренерские указания имеют большее значение. Дело в том, что непосредственно силовое противостояние с противником здесь исключено. Легкоатлет на соревнованиях бежит дистанцию рядом с соперником. Их противостояние сводится к сравнению функциональных возможностей их тел с одной стороны, и тактическому построению бега, противостояние которому со стороны противника хотя и происходит, но косвенно, при отсутствии непосредственного телесного контакта, следовательно, легкоатлет имеет несколько мгновений, а часто и секунд и даже минут на то, чтобы найти тактическое решение в изменившейся ситуации. Главное же – практически все возможные варианты с одной стороны ограничены функциональными возможностями тела, а иные просто не рассматриваются как возможности. Если же такое ограничение не действует, то на уровне мастера спорта все возможные тактические ходы уже известны, продуманы и готовы к использованию.
Таким образом, важно было установить какие аминокислоты преобладают в адаптивных механизмах спортсменов различных спецификаций.
В эсперименте принимали участие легкоатлеты №37, борцы №20 , хоккеисты№22. Показатели сравнивались с контрольной группй студентов, не занимающихся профессионально спртом№15. Исследования проводили в межсоревновательный период, в группах были спортсмены от 1 разряда до мастеров спорта. Для исследования утром натощак забирали кровь из локтевой вены. После центрифугирования плазму депротеинезировали 50% раствором сульфосалициловой кислоты и исследовали на содержание свободных аминокислот на ионообменном анализаторе аминокислот ААА 339М «Микротехника». Уровень кортизола в крови исследовался на иммуноферментном анализаторе.Затем проводили корреляционный анализ количества аминокислот у спортсменов различных спецификаций и уровня мастерства с показателями морфометрии и показателями состояния сердечнососудистой и дыхательной системы, психологическими характеристиками по Спирмену (r).
Достоверность полученных результатов оценивали с помощью непараметрических критериев по Манну-Уитни (критерий «U»).
Исследования на спортсменах были проведены в Екатеринбургском филиале УралГУФК. Выражаем глубокую благодарность всем сотрудникам, принимавшим участие в работе.
2.4 ПОЛУЧЕННЫЕ РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
2.4.1 Содержание аминокислот, имеющих достоверные отличия от контроля и между группами спортсменов
Таблица 2.1 Суммарное количество аминокислот у представителей разных видов спорта количество в мкмоль/л (M+m(min-max)):
| Виды спорта | Количество аминокислот |
| Контроль | 3128,33+845
(1720,91-3676,83) |
| Легкоатлеты | 5103,78**+958
(4144,87-6502,34) |
| Борцы | 3376,17+957
(2565,74-4180,48) |
| Хоккеисты | 3275,17+832
(1519,55-4752,46) |
* Разница с контролем значима на уровне p<0,05.
** Разница с контролем значима на уровне p<0,01.
Из табл. 2.1 видно, что у спортсменов суммарное количество исследованных аминокислот выше, чем в контрольной группе. При этом наибольшее количество аминокислот в плазме крови у легкоатлетов. У борцов и хоккеистов отличие суммарного количества аминокислот от контрольной группы не достоверно.
Таблица 2.2 Относительное содержание аминокислот (%) группы «ДРЕВНИЕ» у представителей разных видов спорта. (M+m (min- max)):
| аспарагиновая | Серин | Глутамновая | Глицин | Аланин | |
| Контрольная | 1,7+ 0,3
(0.4-3.9) |
6,1+0,5
(3.7-8.2) |
3,9+0,5
(0.7-7.1) |
11,5+0,9
(7.9-15.7) |
12,5+1,1
(7.7-17.5) |
| ЛЕГКОАТЛЕТЫ | 2,6**+0,4
(0.8-4.3) |
8,2*+0,7
(6.4-11.8) |
2,6*+0,4
(0.9-4.9) |
12,8+0,5
(10.1-14.4) |
12,5+0,7
(9.5-15.9) |
| БОРЦЫ | 1,4+0,3
(0.3-3.8) |
6,2+0,2
(3.9-6.9) |
6,3*+0,8
(1.9-14.2) |
12,1+0,7
(7.8-15.5) |
12,9+0,9
(7.8-17.3) |
| ХОККЕИСТЫ | 1,0*+0,3
(0.4-1.6) |
5,7+0,6 (2.7-9.5) | 4,2+0,6
(2.9-6.7) |
14,1+1,8 (5.9-21.0) | 13,3+1,7
(6.8-19.8) |
* Разница с контролем значима на уровне p<0.05.
** Разница с контролем значима на уровне p< 0.01.
В плазме крови абсолютное (Табл. 2.3) и относительное (Табл. 2.2) содержание аспарагиновой кислоты у борцов не отличается от таковых в контрольной группе. У легкоатлетов эти величины вдвое превосходят контрольные показатели, а у хоккеистов наоборот, почти вдвое меньше, чем в контрольной группе.
Содержание серина (абсолютное) у легкоатлетов более чем вдвое превосходит контрольный показатель, относительное количество серина у них достоверно выше, чем в контроле. У борцов и хоккеистов никакого отличия в содержании серина ни абсолютного, ни относительного нет.
Что касается глутаминовой кислоты, то ее содержание отличается от контрольных показателей только у борцов – почти вдвое выше и абсолютное, и относительное.
Абсолютное количество глицина выше у представителей всех видов спорта. У легкоатлетов – вдвое, у хоккеистов – достоверно, у борцов – как тенденция. Относительное количество глицина у представителей всех представленных видов спорта от контроля достоверно не отличается.
Почти та же картина, как по глицину наблюдается и относительно аланина. Абсолютное количество аланина в плазме крови спортсменов выше, чем в контроле. У легкоатлетов почти вдвое. У борцов и хоккеистов – недостоверно. Относительное количество аланина у представителей всех рассмотренных видов спорта достоверно не отличается.
Таким образом, в группе древних аминокислот отмечается картина различия в содержании у представителей разных видов спорта. Главным образом это относится к абсолютному содержанию. Тем не менее, повышенное относительное содержание аспарагиновой кислоты характерно для легкоатлетов, пониженное — для хоккеистов, повышенное содержание серина – для легкоатлетов, и повышенное содержание глутаминовой кислоты – для борцов.
Таблица 2.3 Абсолютное содержание аминокислот (мкмоль/л (M+m (min- max)) группы «ДРЕВНИЕ» у представителей разных видов спорта:
| Аспарагиновая | Серин | Глутаминовая | Глицин | Аланин | |
| Контрольная | 58,5+17,3
(14,2-195,2) |
192,2+27,3
(64,0-358,0) |
135,0+29,5
(12,2-318,8) |
366,7+47,9
(126,9-586,1) |
397,5+61,4
(210,6-672,1) |
| ЛЕГКОАТЛЕТЫ | 143,7**+28,4 (23,6-273.5) | 431,7**+70,5 (203.1-753.8) | 123,3+15,1
(60.9-183.3) |
667,9**+68,5 (325.7-984.0) | 625.5*+86,0 (350.7-925.2) |
| БОРЦЫ | 44,2+10,4
(12.0-105.7) |
188,0+22,9
(79.5-333.01) |
231,1*+52,1 (66.6-504.1) | 386,9+47,3 (194.5-658.8) | 406.8+43,6 (217.2-612.9) |
| ХОККЕИСТЫ | 37,1*+10,1
(4.9-103.4) |
194,2+26,1
(71.2-335.5) |
127,3+19,5
(67.1-289.3) |
461,2*+59,7 (184.0-792.9) | 437.4+52,9 (115.4-647.7) |
* Разница с контролем значима на уровне p< 0.05.
** Разница с контролем значима на уровне p<0.01.
Таблица 2.4 Относительное содержание аминокислот производных глутаминовой кислоты (%) у представителей разных видов спорта (M+m (min- max)):
| Глутамин | пролин | орнитин | аргинин | |
| контрольная | 15,8+0,9 (12.4-22.7) | 5,9+0,7 (3.0-10.8) | 3,1+0,6 (1.5-8.6) | 1,9+0,2 (0.8-2.9) |
| ЛЕГКОАТЛЕТЫ | 18,2*+1,4 (15.3-26.2) | 7,4*+1,0 (3.5-12.2) | 2,9+0,3 (2.2-4.9) | 1,9+0,3 (0.2-3.4) |
| БОРЦЫ | 18,4*+1,7 (4.1-22.7) | 5,9+1,8 (0.0-19.7) | 2,2*+0.2 (1.3-3.1) | 2,0+0,2 (1.1-4.3) |
| ХОККЕИСТЫ | 16,7+0,8 (13.4-22.3) | 3,1**+0,7 (1.0-9.1) | 2,8+0,3 (1.9-5.1) | 1,9+0,2 (1.2-3.2) |
* Разница с контролем значима на уровне p<0.05.
** Разница с контролем значима на уровне p<0.01.
Абсолютное количество глутамина в плазме крови (Табл. 2.5) у спортсменов выше, чем в контроле, у легкоатлетов почти вдвое, у борцов и хоккеистов недостоверно. Относительное количество глутамина (Табл. 2.4) у легкоатлетов и борцов достоверно выше, чем в контроле.
Абсолютное содержание пролина у легкоатлетов почти вдвое выше чем в контроле, у борцов не отличается от контроля, у хоккеистов – почти вдвое ниже, чем в контроле. Абсолютно то же самое можно сказать и об относительном содержании пролина (Табл. 2.4) – у легкоатлетов выше, у хоккеистов ниже, чем в контроле, а у борцов – не отличается от такового.
Абсолютное количество орнитина и аргинина в плазме крови легкоатлетов выше, чем в контроле (p<0.05), а у борцов – ниже. У хоккеистов и абсолютное и относительное содержание орнитина в плазме крови не отличаются от таковых в контроле.
Таблица 2.5 Абсолютное содержание (мкмоль/л (M+m(min-max)) аминокислот производных глутамата у представителей разных видов спорта:
| Глутамин | Пролин | Орнитин | Аргинин | |
| Контрольная | 498,1+61,0
(228.0-837.4) |
185,6+27,5
(52.3-330.7) |
99,6+21,4
(42.2-263.1) |
55,7+5,7
(25.0-77.0) |
| ЛЕГКОАТЛЕТЫ | 899,5**+68,3
(672.5-1100.3) |
397,7*+81,5
(104.2-785.7) |
157,0*+29,7
(66.4-314.5) |
102,1**+25,7
(32.02-221.4) |
| БОРЦЫ | 502,6+92,4
(144.8-1088.1) |
161,9+79,1
(54.2-827.2) |
70,8*+6,2
(45.2-108.6) |
75,3*+8,1
(35.0-125.7) |
| ХОККЕИСТЫ | 560,42+78,7
(105.5-1066.6) |
96,41**+27,7
(41.5-318.5) |
95,22+17,8
(24.7-244.6) |
65,84+9,8
(15.8-145.8) |
* Разница с контролем значима на уровне p<0.05.
** Разница с контролем значима на уровне p<0.01.
Таблица 2.6 Относительное содержание серосодержащих аминокислот (%) у представителей разных видов спорта (M+m(min-max)):
| Таурин | Цистин | Метионин | |
| Контрольная | 1,8+0,3
(0.26-3.19) |
2,5+0,3
(0.25-3.92) |
0,9+0,1
(0.4-1.5) |
| ЛЕГКОАТЛЕТЫ | 0,7**+0,2
(0.2-1.5) |
2,2+0,2
(1.3-3.0) |
0,4**+0,1
(0.3-0.6) |
| БОРЦЫ | 0,9**+0,2
(0.3-1.4) |
3,3+0,2
(2.5-4.3) |
0,9+0,1
(0.4-1.8) |
| ХОККЕИСТЫ | 1,7+0,5
(0.4-3.6) |
4,0*+0,5
(2.5-6.1) |
1,1+0,2
(0.2-1.4) |
* Разница с контролем значима на уровне p< 0.05.
** Разница с контролем значима на уровне p<0.01.
Абсолютное содержание таурина в плазме крови у легкоатлетов и борцов ниже, чем в контроле, причем у борцов – вдвое (Табл.2.7). То же самое можно сказать и об относительном содержании таурина (Табл.2.6). У хоккеистов содержание таурина не отличается от контроля.
Абсолютное содержание цистина у представителей всех видов спорта выше, чем в контроле (Табл. 1.7). Относительное же содержание цистина достоверно выше контроля только у хоккеистов, у борцов и легкоатлетов этот показатель не отличаетс он такового в контроле.
Абсолютное содержание метионина в плазме крови у борцов и хоккеистов достоверно выше, чем в контроле, а у легкоатлетов не отличается от такового. У легкоатлетов, таким образом, вполне ожидаемо отмечено снижение вдвое, по сравнению с контролем, содержание метионина. У борцов и хоккеистов относительное содержание метионина не отличается от такового в контроле.
Таблица 2.7 Абсолютное содержание (мкмоль/л (M+m (min-max)) серосодержащих аминокислот у представителей разных видов спорта:
| Таурин | Цистин | Метионин | |
| Контрольная | 60,7+13,6
(40.0-121.1) |
76,4+11,8
(7.6-131.9) |
25,5+2,3
(16.4-43.8) |
| ЛЕГКОАТЛЕТЫ | 41,3*+12,2
(19.1-102.7) |
109,6*+11,5
(57.5-145.0) |
23,1+2,1
(18.01-34.3) |
| БОРЦЫ | 29,2**+3,5
(19.2-47.2) |
110,7*+11,2
(56.6-163.9) |
32,8*+5,0
(15.7-66.7) |
| ХОККЕИСТЫ | 61,9+12,9
(14.0-137.2) |
127,6*+16,7
(48.2-212.5) |
34,1*+8,6
(6.2-62.3) |
* Разница с контролем значима на уровне p<0.05.
** Разница с контролем значима на уровне p<0.01.
Абсолютное содержание валина в плазме крови у борцов и хоккеистов ниже, чем в контроле, а у легкоатлетов – не отличается от контроля. Относительное же содержание валина у хоккеистов, и, особенно, у легкоатлетов ниже, чем в контроле (Табл. 2.8 и 2.9).
Таблица 2.8 Абсолютное содержание (мкмоль/л (M+m(min-max)) разветвленных аминокислот у представителей разных видов спорта:
| Валин | Изолейцин | Лейцин | |
| Контрольная | 234,2+38,3
(106.3-458.5) |
43,9+5,3
(26.7-74.0) |
122,2+14,2
(78.9-206.0) |
| ЛЕГКОАТЛЕТЫ | 234,1+26,7
(141.1-318.6) |
72,4*+8,3
(29.2-95.0) |
158,6*+25,7
(56.0-282.2) |
| БОРЦЫ | 181,3*+32,8
(0.0-407.4) |
56,6*+6,2
(28.0-24.3) |
130,9+11,0
(78.2-195.7) |
| ХОККЕИСТЫ | 177,3*+17,7
(110.8-243.3) |
56,1+9,4
(10.5-101.2) |
121,6+14,3
(68.1-185.1) |
* Разница с контролем значима на уровне p< 0.05.
** Разница с контролем значима на уровне p<0.01.
Абсолютное содержание изолейцина в плазме крови у спортсменов выше, чем в контоле: в полтора раза — у легкоатлетов, достоверно – у борцов и недостоверно – у хоккеистов. Относительное содержание изолейцина у спортсменов не отличается от контроля.
Абсолютное содержание лейцина в плазме крови у легкоатлетов выше контроля, а у борцов и хоккеистов не отличается от такового. Относительное же содержание лейцина все же ниже, чем в контроле именно у легкоатлетов. У борцов и хоккеистов этот показатель не отличается от контроля.
Таблица 2.9 Относительное содержание (%) разветвленных аминокислот у представителей разных видов спорта (M+m (min-max)):
| валин | Изолейцин | Лейцин | |
| Контрольная | 7,5+0,8
(3.8-11.9) |
1,5+0,2
(0.7-3.1) |
4,0+0,3
(2.7-6.0) |
| ЛЕГКОАТЛЕТЫ | 4,6*+0,4
(3.3-6.2) |
1,4+0,1
(0.9-2.0) |
3,0*+0,3
(1.9-4.3) |
| БОРЦЫ | 6,4+1,2
(0.2-15.8) |
1,4+0,1
(1.0-2.8) |
3,5+0,4
(2.9-6.7) |
| ХОККЕИСТЫ | 5,8*+0,4
(3.5-9.1) |
1,6+0,2
(0.6-3.4) |
3,7+0,2
(2.4-4.7) |
* Разница с контролем значима на уровне p<0.05.
** Разница с контролем значима на уровне p<0.01.
Таблица 2.10 Абсолютное содержание (мкмоль/л (M+m(min-max)) циклических аминокислот у представителей разных видов спорта:
| тирозин | фенилаланин | триптофан | гистидин | |
| Контрольная | 55,2+5.5
(36.4-92.7) |
47,1+4.6
(21.2-78.9) |
30,1+8.4
(2,6 -74.9) |
83,3+6.2
(56.0-111.69) |
| ЛЕГКОАТЛЕТЫ | 100,4**+11.1
(47.2-133.8) |
75,5*+6.8
(60.2-113.1) |
75,2*+16.3
(5.6-125.1) |
115,6*+15.1
(74.4-175.5) |
| БОРЦЫ | 54,8+4.0
(32.3-79.6) |
55,7+5.5
(29.3-90.7) |
6,6**+2.5
(0.0-21,6) |
92,4+9.7
(45.6-135.2) |
| ХОККЕИСТЫ | 66,7+7.6
(37.8-111.2) |
59,8*+7.7
(31.5-118.8) |
7,3**+2,8
(0.0-22.8) |
94,2+12.8
(35.0-166.5) |
* Разница с контролем значима на уровне p <0.05.
** Разница с контролем значима на уровне p <0.01.
Абсолютное содержание тирозина (Табл. 2.10) у легкоатлетов почти вдвое превышает таковое в контрольной группе. У борцов и хоккеистов содержание тирозина в плазме крови не отличается от контроля. Тем не менее, у легкоатлетов относительное содержание тирозина (Табл. 2.11) неотличимо от контроля, как и у борцов и хоккеистов.
Абсолютное содержание фенилаланина у спортсменов выше, чем в контрле (Табл. 2.10), достоверно – у хоккеистов и в полтора раза – у легкоатлетов. У борцов – недостоверно. Чего не скажешь об относительном содержании фенилаланина (Табл. 2.11). Относительное содержание фенилаланина в плазме крови у спортсменов не имеет достоверных отличий от такового в контроле.
Особое место занимает триптофан. Его абсолютное количество у легкоатлетов вдвое превышает таковое в контроле, а у борцов и хоккеистов его абсолютное количество более, чем в 4 раза ниже, чем в контроле. Практически то же самое можно сказать об относительном содержании триптофана – у легкоатлетов его, относительно общего количества аминокислот, больше, а у борцов и хоккеистов меньше, чем в контроле.
Абсолютное содержание гистидина у спортсменов выше, чем в контроле, у легкоатлетов – достоверно (p<0,05), тем не менее, у легкоатлетов относительное содержание гистидина ниже, чем в контроле.
Таблица 2.11 Относительное содержание циклических аминокислот (%) в плазме крови у представителей разных видов спорта (M+m (min-max)):
| Тирозин | Фенилаланин | Триптофан | Гистидин | |
| Контроль | 1,8+0,1
(1,3-2,6) |
1,6+0,2
(0,8-3,0) |
0,8+0,2
(0,4-2,0) |
2,8+0,1
(2,0-4,9) |
| ЛЕГКОАТЛЕТЫ | 2,0+0,2
(1,5-3,2) |
1,5+0,1
(1,0-2,0) |
1,4*+0,2
(0,2-2,1) |
2,1*+0,1
(1,6-2,7) |
| Борцы | 1,7+0,1
(1,2-2,4) |
1,8+0,2
(1,1-3,5) |
0,1**+0,0
(0,0-0,6) |
2,9+0,2
(1,7-3,7) |
| ХОККЕИСТЫ | 2,1+0,1
(1,3-3,3) |
1,9+0,2
(1,3-2,7) |
0,2**+0,1
(0,0-0,6) |
2,9+0,2
(1,9-3,9) |
* Разница с контролем значима на уровне p <0.05.
** Разница с контролем значима на уровне p <0.01.
Таким образом установлено, что занятия разными видами спорта по- разному влияют на метаболизм аминокислот. Исследование содержания аминокислот показало преобладание того или иного метаболита у представителей разных видов спорта.
У легкоатлетов суммарное количество аминокислот в плазме крови больше, чем в контроле и у представителей других исследованных видов спорта более, чем в полтора раза. Соответственно и абсолютное количество каждой из аминокислот у легкоатлетов выше, чем в контроле. Исключение составляют глутаминовая кислота, метионин, валин и таурин. Абсолютное количество глутаминовой кислоты, метионина и валина в плазме крови легкоатлетов не отличается от контроля, а количество таурина по сравнению с контролем понижено. Соответственно, относительное содержание этих аминокислот у легкоатлетов так же понижено. Абсолютное содержание лейцина у легкоатлетов повышено, но настолько незначительно, что его относительное содержание все-таки понижено по сравнению с контролем. Обращает на себя внимание резко повышенное содержание аспарагиновой кислоты, серина, глутамина, пролина и триптофана. Содержание этих аминокислот велико не только по абсолютной величине, они выпадают из общего баланса аминокислот — их относительное содержание у легкоатлетов тоже повышено.
Суммарное количество аминокислот у борцов не имеет достоверных отличий от контроля. Достоверное превышение по абсолютному количеству аминокислот у борцов имеют глутаминовая кислота, глутамин, аргинин, цистеин, метионин, валин. Достоверно понижено по сравнению с контролем содержание таурина и триптофана. Но важным является отличие по относительному содержанию. Для борцов характерно повышенное относительное содержание глутаминовой кислоты и глутамина, и пониженное количество орнитина, таурина и триптофана.
Суммарное количество аминокислот в плазме крови хоккеистов минимально, по сравнению с другими исследованными группами. Баланс аминокислот, характерный для хоккеистов, определяется пониженным абсолютным, а, главное, относительным содержанием аспарагиновой кислоты, пролина, валина и триптофана, и повышенным содержанием цистеина.
Таблица 2.12 отклонения от контроля по относительному содержанию аминокислот у представителей разных видов спорта:
| аминокислоты | легкоатлеты | Борцы | Хоккеисты |
| Аспарагиновая | + | — | |
| Серин | + | ||
| Глутаминовая | — | + | |
| Глутамин | + | + | |
| Пролин | + | — | |
| Орнитин | — | ||
| Таурин | — | — | |
| Метионин | — | ||
| Цистеин | + | ||
| Валин | — | — | |
| Триптофан | + | — | — |
| Всего | +5, -4 | +2, -3 | +1, -4 |
Все исследованные спортсмены – люди одного пола, возраста, ведущие одинаковый образ жизни, имеющие сходный род занятий. Различаются они по виду спорта, то есть характер физической нагрузки, и образ действий, связанный с видом спорта, определяют различия в метаболизме. В таблице 2.12 представлены различия по относительному содержанию аминокислот в плазме крови у представителей разных видов спорта. Видно, что наибольшие изменения аминокислотного профиля в плазме крови наблюдается у легкоатлетов, что по, видимому связано, с большей выраженностью реконструктивного протеолиза. У борцов и хоккеистов суммарное количество аминокислот не отличается от контроля, тем не менее, профиль аминокислот у представителей этих двух видов спорта имеет отчетливые различия. Это выражается в большем количестве глутаминовой кислоты и глутамина у борцов и цистеина у хоккеистов, и меньшем количестве орнитина и таурина у борцов и аспарагиновой кислоты и валина у хоккеистов. Объединяет же борцов и хоккеистов сниженное относительное содержание триптофана, в отличие от легкоатлетов, у которых количество триптофана повышено.
Спортсмены на момент исследования неоднородны по степени достижения успеха в своей деятельности. Успешность деятельности в спорте определяется системой присвоения спортивных разрядов. В нашем исследовании принимали участие спортсмены, имеющие квалификацию 1разряда, кандидаты в мастера спорта, мастера спорта и мастера спорта международного класса. Следующая глава посвящена исследованию аминокислотного состава плазмы крови в зависимости от уровня мастерства спортсменов.
2.4.2 Содержание аминокислот, имеющих достоверные отличия от контроля и между группами в плазме крови спортсменов в зависимости от уровня их мастерства
Общее количество исследуемых аминокислот у представителей разных видов спорта представлено в таблице 2.13.
В табл.2.13 видно, что у легкоатлетов в среднем количество аминокислот в плазме крови заметно (более чем в полтора раза) превышает этот показатель в контроле. При этом у перворазрядников эта величина больше чем в контроле более, чем вдвое. Затем, по мере роста квалификации общее количество аминокислот в плазме крови легкоатлетов снижается и у МСМК эта величина уже ниже, чем в контроле, хотя не достоверно, но зато достоверно ниже, чем у МС.
Таблица 2.13 Суммарное количество аминокислот в плазме крови у представителей разных видов спорта количество в мкмоль/л (M+m (min-max)):
| легкоатлеты | Борцы | Хоккей | |||||||
| Среднее | N | + | Среднее | N | + | Среднее | N | + | |
| 1р | 6419,53** | 10 | 523 | 3665,48* | 17 | 576 | |||
| КМС | 5944,94** | 12 | 448 | 3986,56* | 10 | 413 | 3125,20 | 17 | 534 |
| МС | 4259,90* | 10 | 115 | 2861,42 | 10 | 356 | 3034,78 | 18 | 985 |
| МСМК | 2952,36 | 5 | 387 | ||||||
| Среднее | 5103,79** | 37 | 520 | 3198,96 | 20 | 316 | 3247,89 | 22 | 360 |
| Контроль | 3128,33 | 27 | 388 | 3128,33 | 27 | 388 | 3128,33 | 27 | 388 |
* Разница с контролем значима на уровне p<0,05.
** Разница с контролем значима на уровне p<0,01.
У борцов общее количество аминокислот в плазме крови в среднем не отличается от контроля. При этом у КМС эта величина больше, чем в контроле и чем у МС. Налицо та же тенденция, что и у легкоатлетов – снижение общего содержания аминокислот в плазме крови с ростом мастерства.
Схема 2. Динамика кортизола и суммы свободных аминокислот как показателей хронического стресса в крови у спортсменов:

У хоккеистов и среднее количество аминокислот в плазме крови и эта величина у представителей разного уровня квалификации не имеет достоверных отличий от контроля, за исключением перворазрядников, у которых суммарное количество аминокислот выше, чем в контроле (p<0,05). Тенденция к снижению количества аминокислот с ростом квалификации у хоккеистов вполне отчетливо видна (Табл.2.13). Суммарное количество аминокислот в определенной степени отражает уровень стрессового состояния у спортсмена («реконструктивный протеолиз») [197,260]. Как видим его уровень выше и дольше сохраняется у легкоатлетов, по сравнению с борцами и хоккеистами. Это подтверждается динамикой количества глюкокортикоидов у исследуемых спортсменов. (Схема 2.2)
Представленная динамика суммы аминокислот соответствует изменениям уровня глюкокортикоидов в крови исследуемых спортсменов, то есть уровню хронческого стрессового воздействия занятий спортом на организм. Так максимальное количество кортизола мы отмечаем у легкоатлетоа с 1го разряда до КМС, то есть становление легкоатлета требует субъективно больших усилий по сравнению с борцами и легкоатлетами. Снижение уровня суммы свободных аминокислот и, соответственно, кортизола мы наблюдаем только у спортсменов на уровне МСМК. У борцов и хоккеистов сумма свободных амрнокислот на уровне 1разряд — КМС повышается гораздо в меньшей степени, чем у легкоатлетов, а уже на уровне МС отмечается снижение суммы аминокислот ниже уровня контроля, равно как и кортизола. Таким образом, есть основания полагать, что исследованные борцы и хоккеисты более стрессустойчивы, чем легкоатлеты. Что находит свое отражение и в особенностях метаболизма аминокислот.
Легкоатлеты
Таблица 2.14 Абсолютное содержание (мкмоль/л) «древних» аминокислот в крови у легкоатлетов в зависимости от квалификации (M+m(min-max)):
| Квалиф | Аспарагиновая | Серин | Глутаминовая | Глицин | Аланин |
| Контрольная | 58,5+17,3
(14.2-195.2) |
192,2+27,3
(64,0-358,0) |
135,0+29,5
(12,2-318,8) |
366,7+47,9
(126,9-586,1) |
397,5+61,4
(210,4-672,1) |
| 1разряд | 179,1*+21 (68.1-322.8) | 569,4*+128,9
(451.1-806.4) |
60,9*+15,2
(18.4-203.8) |
925,0*+119,5
(752.2-1108.4) |
611,1*+-78,4(315,6-873,2 |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 183,5*+40,3 (119.0-273.5) | 544,4*+181.7 (427.2-753.8) | 107,2+10,2 (107,8-183,3) | 800,5*+105.3 (699.3-909.6) | 761,3**+-79,7)389.1-944,6) |
| МАСТЕРА СПОРТА | 126,5*+4,8 (73.0-229.9) | 308,2*+58,4 (166.8-449.6) | 147,3+28,0 (60.2-246.3) | 498,8+49,5
(310.0-621.0) |
507,0+-68,8(308,2-769,4) |
| МАСТЕРА СПОРТА МЕЖДУНАРОДНОГО КЛАССА | 23,6*+4,1
(16.9-42.5) |
203,1+29.0
(91.2-251.4) |
146,1+16,8 (96.7-143.6) | 50,7**+9.6
(18.4-76.5) |
470,6+-75,7(276,8-615,5) |
* Разница с контролем значима на уровне p <0.05.
** Разница с контролем значима на уровне p <0.01.
Абсолютное и относительное количество аспарагиновой кислоты в плазме крови у легкоатлетов выше, чем в контроле (Табл.2.14). У перворазрядников и КМС абсолютное количество аспарагиновой кислоты увеличено в 3 раза. Далее, с ростом мастерства абсолютное количество аспарагиновой кислоты в плазме крови легкоатлетов снижается, и на уровне МСМК вдвое ниже, чем в контроле. Практически то же самое можно сказать об относительном количестве аспарагиновой кислоты. С уровня мастера спорта он начинает снижаться и на уровне МСМК снижен более, чем в 2 раза по сравнению с контролем.
Динамика содержания глутаминовой кислоты в плазме крови легкоатлетов иная. На уровне первого разряда абсолютное количество её резко понижено (более, чем в 2 раза), затем, с ростом мастерства повышается до уровня контроля. Изменения относительного количества глутаминовой кислоты с ростом мастерства аналогично абсолютному, но колебания несколько резче, чем абсолютного количества.
Обращает на себя внимание, что количество всех древних аминокислот на уровне 1 разряда и КМС увеличено, а на уровне МСМК абсолютное количество серина и глутаминовой кислоты выравнивается с контролем, а абсолютное количество аспарагиновой кислоты и глицина даже резко снижено по сравнению с контролем – в два раза для аспарагиновой кислоты и в семь раз для глицина.
Таблица 2.15 Относительное содержание (%) древних аминокислот в крови у легкоатлетов в зависимости от квалификации (M+m(min-max)):
| Квалиф | Аспарагиновая | Серин | Глутаминовая | Глицин | Аланин |
| Контроль | 1,7+ 0,3
(0,4-3,9) |
6,1+0,5
(3,7-8,2) |
3,9+0,5
(0,7-7,1) |
11,5+0,9
(7,9-15,7) |
12,5+1,1
(7,7-17,5) |
| 1разряд | 2,7*+1,1
(0.8-3.4) |
8,8+1,9 (4,2-15,1) | 0,9*+0,1
(0.3-0.9) |
14,4*+1,9 (8.5- 16.0) | 27,0**+1,2 (9.5-31.2) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 3,1*+1,2
(1.83-4.3) |
9,1*+2,6 (6.57-11.8) | 2,4*+0,3
(2.1-2.8) |
13,5+1,1 (12.1-14.3) | 28,2**+2,8 (11.9-33.6) |
| МАСТЕРА СПОРТА | 2,9*+0,8
(0.6-2.9) |
7,2+1,0 (6.4-7.9) | 2,5+0,3
(2.2-2.7) |
11,6+2,1 (10.1-13.1) | 24,3**+4,1 (11.0-32.7) |
| МАСТЕРА СПОРТА МЕЖДУНАРОДНОГО КЛАССА | 0,8*+0,2
(0.2-1.4) |
6,8+1,2
(2.8-11.6) |
4,9*+0,6
(1.2-8.9) |
11,8+3.1
(8.4-16.8) |
24,5**+2,8
(10.5-26.5) |
* Разница с контролем значима на уровне p<0.05.
** Разница с контролем значима на уровне p<0.01.
Из представленных таблиц видно, что у всех легкоатлетов в крови отмечается повышенное количество аланина. По относительным значениям примерно в 2 раза (Табл.2.15). А по абсолютным показателям это превышение особенно заметно в группах 1 разрядников и КМС (достоверно отличающиеся от контроля результаты). Тенденденция к превышению количества аланина наблюдается и в группах МС и МСМК, но это превышение уже статистически значимо от контроля не отличается.
Аланин это одна из «древних» аминокислот в которую метаболизируется молочная кислота, образующаяся в мышцах при повышенных физических нагрузках. И судя по тому, что у легкоатлетов во всех группах (особенно у начинающих спортсменов) повышено количество аланина, видимо, это является следствием повышенного образования в мышцах молочной кислоты. Чего кстати не наблюдается у других рассматриваемых нами групп спортсменов-борцов и хоккеистов, что видимо связано с другим характером нагрузок и другими адаптивными возможностями легкоатлетов по сравнению с борцами и хоккеистами.
Общее абсолютное количество древних аминокислот у легкоатлетов с ростом мастерства уменьшается за счет АСП, СЕР, ГЛИ. А вот абсолютное и относительное количество ГЛУ у легкоатлетов с ростом мастерства многократно возрастает.
Таблица 2.16 Абсолютное содежание (мкмоль/л (M+m(min-max)) серосодержащих аминокислот в крови у легкоатлетов в зависимости от квалификации:
| Квалиф | Таурин | Цистеин | Метионин |
| Контроль | 60,7+13.6
(4.0-121.1) |
76,4+11.8
(7.6-131.9) |
25,5+2.3
(16.4-43.8) |
| 1разряд | 34,0**+11,2
(6.8-54.9) |
120,6*+26,5
(45.1-241.6) |
19,2+4.3
(4,8-36,2) |
| КАНДИДАТЫ В МАСТЕРА | 46,2+19.2
(11.9-102.7) |
119,3*+18.7
(85.2-145.0) |
27,0+7.4
(19.5-34.3) |
| МАСТЕРА СПОРТА | 53,70+3.2
(51.3-56.0) |
115,58*+12.8
(99.4-131.7) |
21,70+0.3
(21.4-21.9) |
| МАСТЕРА СПОРТА МЕЖДУНАРОДНОГО КЛАССА | 9,1**+3,1
(5,8-14,6) |
57,5+12,8
(28,6-78,4) |
18,0+5,12
(8.9-26.1.20 |
* Разница с контролем значима на уровне p <0.05.
** Разница с контролем значима на уровне p <0.01.
Содержание серосодержащих аминокислот в плазме крови у легкоатлетов представлены в таблицах 2.15 (абсолютное) и 2.16 (относительное). Абсолютное и отностительное содержание таурина у легкоатлетов значительно ниже, чем в контрольной группе (Табл.2.16, 2.17). У перворазрядников абсолютное и относительное содержание таурина в два и три раза соответственно ниже, чем в контроле. С ростом мастерства абсолютное и относительное содержание таурина возрастает почти до уровня контроля, и резко снижается у МСМК.
Абсолютное содержание цистеина в плазме крови у легкоатлетов выше, чем в контроле, а относительное не отличается от контроля (Табл.2.16, 2.17). Максимальное абсолютное содержание цистеина у легкоатлетов наблюдается у перворазрядников, а минимальное – у МСМК.
Абсолютное количество метионина с ростом мастерства достоверне не изменяется. Относительное количество метионина с началом занятий легкой атлетикой резко падает, но затем, по мере роста мастерства возрастает почти до уровня контрольной группы.
Таблица 2.17 Относительное содержание (%) серосодержащих аминокислот в крови у легкоатлетов в зависимости от квалификации (M+m (min-max)):
| Квалиф | Таурин | Цистеин | Метионин |
| Контроль | 1,8+0,3 (0.5-4.1) | 2,5+-0,5 (0.8-3.5) | 0,9+0,2 (0.0-1.9) |
| 1разряд | 0,5**+0,1
(0.0-1.2) |
1,8*+0,4
(0.3-3.8) |
0,3*+0,1
(0.0-0.8) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 0,7*+0.1 (0.2-1.5) | 2,0+0.6 (1.3-2.5) | 0,4*+0.1 (0.3-0.5) |
| МАСТЕРА СПОРТА | 1,2*+0.2 (1.2-1.2) | 2,7+0.4 (2.4-3.0) | 0,5+0.2 (0.4-0.5) |
| МАСТЕРА СПОРТА МЕЖДУНАРОДНОГО КЛАССА | 0,3**+0,1
(0.0- 1,1) |
1,9+0,3
(0.2-2.7) |
0,6+0,3
(0.0- 2.4) |
* Разница с контролем значима на уровне p<0.05.
** Разница с контролем значима на уровне p<0.01.
Общее количество серосодержащих аминокислот у легкоатлетов возрастает до уровня МС, затем снижается. Это относится к ТАУ и ЦИС. МЕТ с ростом мастерства возрастает монотонно.
Таблица 2.18 Абсолютное содержание (мкмоль/л (M+m(min-max)) аминокислот производных глутаминовой кислоты в крови у легкоатлетов в зависимости от квалификации:
| Квалиф | Глутамин | Пролин | Орнитин | Аргинин |
| Контроль | 498,1+61,0 (228.0-837.4) | 185,6+27,5 (52.3-330.7) | 99,6+21,4 (42.2-263.1) | 55,7+5,7 (25.0-77.0) |
| 1разряд | 1100,3**+254,8
(531.8-1987.1) |
785,7**+108,5
(584.2-912.8) |
314,5**+55,4
(210.0-431.8) |
221,4**+42,8
(46.5-365.9) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 979,0**+76,0 (776.3-1094.1) | 390,3**+34,6 (277.3-524.1) | 156,1*+14,2 (130.7-189.5) | 112,1**+29,6 (89.0-158.0) |
| МАСТЕРА СПОРТА | 742,6*+49,0 (672.5-812.6) | 361,5*+34,5 (294.6-428.3) | 124,9+11,5 (111.1-138.8) | 49,1+12,4 (12.0-86.2) |
| МАСТЕРА СПОРТА МЕЖДУНАРОДНОГО КЛАССА | 774,40*+96,4
(537.8-962.5) |
104,22*+34,1
(51,4-220.0) |
66,43*+9,8
(36.1-82,6) |
58,75+8,2
(30.1-81,6) |
* Разница с контролем значима на уровне p<0.05.
** Разница с контролем значима на уровне p<0.01.
Содержание аминокислот производных глутаминовой кислоты в плазме крови у легкоатлетов представлены на таблицах 2.17 и 2.18. Абсолютное количество глутамина в плазме крови у легкоатлетов почти вдвое превосходит таковое в контроле (таблица 2.15). Максимум – у перворазрядников, с ростом мастерства происходит снижение количества глутамина, но и на уровне МС и МСМК содержание глутамина превосходит контроль в полтора раза. Обращает на себя внимание то, что относительное содержание глутамина в плазме крови легкоатлетов не имеет достоверных отличий от контроля, то есть увеличение количества глутамина происходит одновременно с увеличеинем суммарного количества аминокислот. На этом фоне относительное количество глутамина у МСМК остается резко повышенным.
По абсолютному и относительному содержанию пролина в плазме крови легкоатлеты, несомненно, являются чемпионами среды представленных видов спорта (Табл. 2.14, 2.15). Среди легкоатлетов же чемпионами по содержанию пролина в плазме крови являются перворазрядники, далее, с ростом мастерства, уровень пролина снижается, достигая минимума у МСМК. У МСМК уровень пролина вдвое ниже, чем в контроле.
Практически то, что сказано о пролине можно сказать об орнитине и аргинине. Многократно по сравнению с контролем их количество повышено у перворазрядников, и в дальнейшем с ростом мастерства снижается.
Таблица 2.19 Относительное содержание (%) аминокислот производных глутаминовой кислоты в крови у легкоатлетов в зависимости от квалификации (M+m(min-max)):
| Квалиф | Глутамин | Пролин | орнитин | Аргинин |
| Контроль | 15,8+0.9 (12.4-22.7) | 5,9+0.7 (3.0-10.8) | 3,1+0.6 (1.5-8.6) | 1,9+0,2 (0.8-2.9) |
| 1разряд | 17,1+0,3
(14.2-22.1) |
12,2**+2.5
(5.0-18.4) |
4,9*+0.6
(2.1-6.8) |
3,4*+0,4
(1.9-5.2) |
| КАДИДАТЫ В МАСТЕРА СПОРТА | 16,4+0.7 (15.6-17.2) | 6,4+0.3 (5.5-8.0) | 2,6+0.3 (2.2-2.9) | 1,8+0.5 (1.4-4.4) |
| МАСТЕРА СПОРТА | 17,4+0.9 (15.3-19.6) | 7,4+0.8 (7.1-9.7) | 2,9+0.5 (2.5-3.3) | 1,1+1.1 (0.2-1.9) |
| МАСТЕРА СПОРТА МЕЖДУНАРОДНОГО КЛАССА | 26,2**+3,2
(18.8-42.6) |
3,5*+0,3
(2.4-5.9) |
2,2+0,6
(1.4-4.2) |
1,9+0,3
(0.8-3.1) |
* Разница с контролем значима на уровне p<0.05.
** Разница с контролем значима на уровне p<0.01.
Таблица 2.20 Абсолютное содержание (мкмоль/л (M+m(min-max)) разветвленных аминокислот в крови у легкоатлетов в зависимости от квалификации:
| Квалиф | Валин | Изолейцин | Лейцин |
| Контроль | 236,2+38.3
(106.3-458.5) |
43,9+5.3
(26.7-74.0) |
122,2+14.2
(78.9-206.0) |
| 1разряд | 212,4+31.7
(214.8-376.8) |
95,0**+10,6
(50.7-204.1) |
141,2+18,9
(83.6-223,1) |
| КАДИДАТЫ В МАСТЕРА СПОРТА | 293,8+28.4
(262.7-318.6) |
76,9*+13.2
(63.0-89.4) |
203,4*+38.4
(159.0-282.2) |
| МАСТЕРА СПОРТА | 201,9+21.7
(144.8-259.0) |
76,0**+2.7
(69.1-82.8) |
151,4+15.2
(122.9-179.8) |
| МАСТЕРА СПОРТА МЕЖДУНАРОДНОГО КЛАССА | 141,1*+20,2
(88,4-251,7) |
29,2*+3,6
(16,9-41,5) |
56,0*+8,6
(349-68.3) |
* Разница с контролем значима на уровне p< 0.05.
** Разница с контролем значима на уровне p<0.01.
Абсолютное и относительное количество разветвленных аминокислот в плазме крови легкоатлетов в зависимости от уровня мастерства представлено в таблицах 2.19 и 2.20.
Абсолютное количество валина в плазме крови у легкоатлетов не отличается от такового в контроле (Табл.1.8). Тем не менее, абсолютное количество валина имеет отрицательную динамику, то есть, начиная с уровня кандидата в мастера спорта, эта величина уменьшается, и на уровне МСМК становится в полтора раза ниже контроля. Относительное содержание валина у легкоатлетов возрастает от перворазрядников до кандидатов в мастера спорта, а затем снижается.
Абсолютное количество изолейцина у легкоатлетов выше, чем в контроле и чем у представителей других видов спорта. У перворазрядников его количество вдвое выше, чем в контроле, далее с ростом мастерства уровень изолейцина снижается и на уровне МСМК уже почти на треть ниже, чем в контроле.
Абсолютное количество лейцина в плазме крови у легкоатлетов выше, чем в других группах (Табл. 1.8). У мастеров спорта вдвое выше, чем в контроле, а у мастеров международного класса вдвое ниже, чем в контроле, то есть динамика содержания лейцина аналогична динамикое содержания большинства рассмотренных аминокислот. Примерно такова же динамика относительного содержания лейцина у легкоатлетов с ростом мастерства.
В общем, разветвленные аминокислоты к МСМК снижаются ниже нормы.
Таблица 2.21 Относительное содержание (%) разветвленных аминокислот в крови у легкоатлетов в зависимости от квалификации (M+m(min-max)):
| Квалиф ЛА | Валин | Изолейцин | Лейцин |
| Контроль | 7,5+0.8
(3.8-11.9) |
1,5+0.2 (0.7-3.1) | 4,0+0.3
(2.7-6.0) |
| 1разряд | 3,3*+0,6
(0.9-5.2) |
1,4+0,4
(0,8-2,6) |
2,2*+0,3
(0.9-3.1) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 5,0*+0.5 (4.1-6.0) | 1,3+0.4 (0.9-1.8) | 3,4+-0.6 (2.5-4.3) |
| МАСТЕРА СПОРТА | 4,7*+2.0 (3.3-6.2) | 1,7+0.2 (1.5-2.5) | 3,5+0,4 (2.8-4.3) |
| МАСТЕРА СПОРТА МЕЖДУНАРОДНОГО КЛАССА | 4,7*+2,1
(1,6-6.2) |
0,9*+0.2
(0.3-1.7) |
1,9*+0.3
(0.6-2.8) |
* Разница с контролем значима на уровне p<0.05.
** Разница с контролем значима на уровне p<0.01.
Таблица 2.22 Абсолютное содержание (мкмоль/л (M+m(min-max)) циклических аминокислот в крови у легкоатлетов в зависимости от квалификации:
| Квалиф | Тирозин | Фенилаланин | Триптофан | Гистидин |
| Контроль | 55,2+5.5 (36.4-92.7) | 47,1+4.6 (21.2-78.9) | 30,1+8.4 (4.8-74.9) | 83,3+6.2 (56.0-111.7) |
| 1разряд | 97,5*+11,5
(45.1-152.4) |
75,7*+8,6
(38.9-114.7) |
125,1**+21.0
(68.4-197.9) |
157,9*+28.6
(67.3-218.8) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 112,0**+19.4 (91.4-130.0) | 86,6*+23.8 (66.8-113.1) | 80,7**+40.9 (43.8-124.8) | 124,5*+44.6 (92.9-175.5) |
| МАСТЕРА СПОРТА | 111,1**+12.1 (88.3-133.8) | 66,2*+6.4 (41.7-80.8) | 76,7*+20.1 (32.5-91.0) | 84,2+10.9 (56.5-102.0) |
| МАСТЕРА СПОРТА МЕЖДУНАРОДНОГО КЛАССА | 47,24+6.1
(25.8-74.2) |
60,2+8.5
(29.6-81.4) |
5,6**+1,2
(2.8-8.7) |
74,4+9,6
(38.1-100.5) |
* Разница с контролем значима на уровне p <0.05.
** Разница с контролем значима на уровне p <0.01.
Содержание циклических аминокислот в плазме крови у легкоатлетов отражено в таблице 2.21 (абсолютное содержание), и таблица 2.22 (относительное содержание).
Абсолютное содержание тирозина в плазме крови у легкоатлетов выше, чем в контроле. Уже на уровне первого разряда, уровень тирозина почти вдвое выше, чем в контроле, далее, содержание тирозина еще возрастает, а у МСМК возвращается к норме. Относительное содержание тирозина не отличается от контроля.
Абсолютное содержание фенилаланина у легкоатлетов выше, чем в контроле (таблица №20). Высоким оно остается в каждой квалификационной группе, с тенденией к снижению от КМС к МСМК (Табл. 2.21). При этом относительное содержание фенилаланина в плазме крови легкоатлетов остается неотличимым от контроля независимо от уровня квалификации спортсмена.
Абсолютное содержание триптофана в плазме крови легкоатлетов более, чем вдвое превышает таковое в контроле, что резко выделяет легкую атлетику среди других исследованных нами видов спорта, борцов и хоккеистов, у которых абсолютное содержание триптофана более чем вчетверо ниже, чем в контроле (Табл. 2.20). С ростом квалификации абсолютное количество триптофана у легкоатлетов неуклонно снижается, и на уровне МСМК более чем в пять раз ниже контроля. Динамика относительного содержания триптофана у легкоатлетов аналогична (Табл. 2.23).
Динамика абсолютного и относительного содержания гистидина в плазме крови легкоатлетов аналогична таковой для триптофана.
Таблица 2.23 Относительное содержание (%) циклических аминокислот в крови у легкоатлетов в зависимости от квалификации (M+m(min-max)):
| Квалиф | Тирозин | фениаланин | триптофан | Гистидин |
| Контроль | 1,8+0,1
(1,3-2,6) |
1,6+0,2
(0,8-3,0) |
0,8+0,2
(0,4-2,0) |
2,8+0,2
(2,0-4,9) |
| 1разряд | 1,5+0,3
(0.5-3.6) |
1,2+0,2
(0.5-2.2) |
1,9*+0,3
(1.0-3.6) |
2,4+0,4
(0.9-5.2) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 1,8+0.2 (1.4-2.0) | 1,4+0.3 (1.0-1.7) | 1,3+0.4 (0.6-1.9) | 2,0+0.2 (1.6-2.7) |
| МАСТЕРА СПОРТА | 2,6+0.3 (0.9-3.2) | 1,5+0.2 (1.4-1.7) | 1,7+0.4 (1.5-2.0) | 1,9*+0.3 (1.7-2.2) |
| МАСТЕРА СПОРТА МЕЖДУНАРОДНОГО КЛАССА | 1,6+0,3
(0.8-3.2) |
2,0+0.3
(1.2-3.0) |
0,2*+0.3
(0.0-0.8) |
2,5+0,4
(1.1-4.8) |
* Разница с контролем значима на уровне p <0.05.
** Разница с контролем значима на уровне p <0.01.
Таким образом, на таблице 2.23 мы видим, что суммарное количество аминокислот у легкоатлетов с ростом мастерства снижается, и на уровне МСМК становится достоверно ниже контроля. То есть уровень реконструктивного протеолиза у легкоатлетов очень высок на уровне первого разряда и ниже контроля на уровне высочайшего мастерства. Здесь надо отметить, что первый разряд – это не новички. Для того, чтобы стать перворазрядником спортсменам надо интенсивно тренироваться на протяжении 3-4 и более лет. Мощный реконструктивный протеолиз является пропуском в мир высоких достижений в легкой атлетике. Достигнув же высокого мастерства, спортсмены уже не нуждаются в формировании структурного следа адаптации, так как тело уже сформировано, что находит свое выражение в снижении интенсивности метаболизма аминокислот.
Что касается абсолютного количества каждой из аминокислот в плазме крови у легкоатлетов, то с ростом мастерства либо неуклонно понижается, достигая минимума у МСМК, либо достигает максимума на уровне КМС или МС, после чего опять же достигает минимума на уровне МСМК. Это вполне согласуется с динамикой суммы аминокислот. Из этого ряда выпадает одна аминокислота – глутаминовая. Ее абсолютное количество с ростом мастерства возрастает, и достигает максимума на уровне МСМК. Поэтому есть смысл обратить внимание на обмен глутаминовой кислоты как на возможный маркер мастерства, или возможности достижения мастерства.
С ростом мастерства у легкоатлетов не только снижается суммарное количество аминокислот и снижение количества отдельных аминокислот (за исключением глутаминовой), но и происходит изменение соотношения аминокислот между собой. Так относительное количество аспарагиновой, глутаминовой кислот, метионина, глутамина, валина и фенилаланина возрастает, относительное количество глицина, таурина, пролина, орнитина, аргинина и изолейцина снижается, а относительное содержание серина, цистеина, лейцина, тирозина, триптофана и гистидина не изменяется.
Борцы
Изменений количества аланина у борцов ни по абсолютным ни по относительным значениям в крови не отмечено поэтому в таблицах его значение не приводится.
На таблице 2.24 представлена динамика содержания абсолютного количества «древних» аминокислот в плазме крови у борцов в зависимости от уровня мастерства, а на таблице 2.25 динамика относительного количества древних аминокислот. Абсолютное и относительное количество аспарагиновой кислоты в плазме крови у борцов ниже, чем в контроле, при этом с ростом мастерства от КМС до МС возрастает до уровня контроля.
Количество серина и глицина как абсолютное, так и относительное у борцов с ростом мастерства тоже снижается. То же можно сказать и о глутаминовой кислоте, но надо отметить, что если серин и глицин снижаются до уровня контроля, то абсолютное поличсство глутаминовой кислоты, снижаясь, на уровне мастера спорта остается почти вдвое выше контроля, а ее относительное количество даже не снижается.
Таблица 2.24 Абсолютное содержание (мкмоль/л (M+m(min-max)) группы «древних» аминокислот в крови у борцов:
| Квалиф | Аспарагиновая | Серин | Глутаминовая | Глицин |
| Контрольная | 58,5+17,3
(14.2-195.2) |
192,2+27,3
(64.0-358.0) |
135,0+29,5
(12.2-318.8) |
366,7+47,9
(126.9-586.1) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 18,6*+6,5
(12,0-31,6) |
253,3*+39,8
(213.4-333.0) |
273,6*+89.4
(66.6-485.3) |
559,7*+51,0
(488.8-658.8) |
| МАСТЕРА СПОРТА | 55,1+12,6
(22.3-105.7) |
160,1+21,6
(79.5-231.9) |
232,8*+56.6
(81,4-404,1) |
312,8+38,2
(164.5-467.2) |
* Разница с контролем значима на уровне p<0.05.
** Разница с контролем значима на уровне p<0.01.
Таблица 2.25 Относительное (%) содержание «древних» аминокислот в крови у борцов в зависимости от квалификации (M+m(min-max)):
| Квалиф | аспарагиновая | серин | глутаминовая | Глицин |
| Контрольная | 1,7+0,3
(0.4-3.9) |
6,1+0,5
(0.7-7.1) |
3,9+0,5
(3.7-8.2) |
11,5+0,9
(7.9-15.7) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 0,4*+0,1
(0.3-0.6) |
6,3+0,3
(5.6-6.9) |
6,4*+1.1
(3.9-10.1) |
14,1*+0,7
(13.0-15.5) |
| МАСТЕРА СПОРТА | 1,9+0,3
(1.0-3.8) |
5,6+0,3
(3.9-6.8) |
6,7*+1,3
(3.1-14.2) |
11,1+0,7
(7.8-13.6) |
* Разница с контролем значима на уровне p<0.05.
** Разница с контролем значима на уровне p<0.01.
Таблица 2.26 Абсолютное содержание (мкмоль/л (M+m(min-max)) серосодержащих аминокислот у борцов в зависимости от квалификации:
| Квалиф | Таурин | цистеин | Метионин |
| Контрольная | 60,7+13,6
(4.0-121.1) |
76,4+11,8
(47.6-131.9) |
25,5+2,3
(16.4-43.8) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 35,2*+6.7
(28.0-41.2) |
142,2*+19.0
(127.9-177.1) |
33,4+5.4
(15.7-44.5) |
| МАСТЕРА СПОРТА | 26,6*+12.2
(9.2-47.2) |
97,2+32.7
(56.3-154.9) |
32,5+17.4
(19.5-66.7) |
* Разница с контролем значима на уровне p<0.05.
** Разница с контролем значима на уровне p<0.01.
Содержание серосодержащих аминокислот в плазме крови у борцов представлено на таблице 2.26 (абсолютное) и на таблице 2.27 (относительное).
Абсолютное количество таурина у борцов вдвое ниже, чем в контроле, а с ростом мастерства от КМС до МС имеется тенденция к дополнительному снижению. Относительное количество таурина также у борцов вдвое ниже, чем в контроле, но с ростом мастерства таким же и остается, снижаясь вместе со снижением общего количества аминокислот.
Абсолютное количество цистеина у борцов в полтора раза выше, чем в контроле, а с ростом мастерства происходит снижение цистеина, у мастеров спорта он ниже, чем у кандидатов. Относительное же содержание цистеина в плазме крови у борцов выше, чем в контроле, и с ростом мастерства достоверно не снижается.
Метионина в плазме крови борцов больше, чем в контроле почти в полтора раза. С ростом мастерства от КМС до МС абсолютное количество метионина не меняется. Относительное же количество метионина у борцов не отличается от контрольного, и ростом мастерства не меняется.
Общее абсолютное количество серосодержащих аминокислот у борцов с ростом квалификации уменьшается, как суммарно, так и в отношении каждой из аминокислот.
Общее относительное количество серосодержащих аминокислот в крови у борцов с ростом мастерства изменяется недостоверно.
Таблица 2.27 Относительное содержание (%) серосодержащих аминокислот в крови у борцов в зависимости от квалификации (M+m(min-max)):
| Квалиф | Таурин | цистеин | Метионин |
| Контрольная | 1,8+0,3
(0.5-4.1) |
2,5+0,5
(0.8-3.5) |
0,9+0,2
(0.0-1.9) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 0,8*+0,1
(0.7-1.1) |
3,6*+0,8
(2.6-4.3) |
0,8+0,3
(0.4-1.1) |
| МАСТЕРА СПОРТА | 0,9*+0,3
(0.3-1.4) |
3,4+0,6
(2.5-4.3) |
1,1+0,3
(0.7-1.8) |
* Разница с контролем значима на уровне p<0.05.
** Разница с контролем значима на уровне p<0.01.
Таблица 2.28 Абсолютное содержание (мкмоль/л (M+m(min-max)) аминокислот производных глутаминовой в крови у борцов в зависимости от квалификации:
| Квалиф | Глутамин | Пролин | Орнитин | Аргинин |
| Контрольная | 498,1+61,0
(228.0-837.4) |
185,6+27,5
(52.3-330.7) |
99,6+11,4
(42.2-263.1) |
55,7+5,7
(25.0-77.0) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 773,6*+178,1
(560.0-1088.1) |
158,8+61,0
(35.8-230.9) |
90,0+12,4
(65.1-108.6) |
83,8*+5.7
(72.3-90.5) |
| МАСТЕРА СПОРТА | 386,5+123,6
(144.8-789.6) |
163,2+54,4
(49,5-327.2) |
62,6*+4,5
(45.2-77.8) |
71,7+11,1
(35.0-125.7) |
* Разница с контролем значима на уровне p<0.05.
** Разница с контролем значима на уровне p<0.01.
Содержание аминокислот, производных от глутаминовой в плазме кровни у борцов представлены в таблице 2.28 (абсолютное) и 2.29 (относительное).
Абсолютное содержание глутамина у борцов не отличается от такового в контроле, при этом у кандидатов в мастера абсолютное содержание глутамина достоверно выше, чем в контроле, и, соответственно, у мастеров спорта ниже, чем в контроле – в виде тенденции. Относительное содержание глутамина в плазме крови у борцов имеет аналогичную динамику.
Как абсолютное, так и относительное содержание пролина в плазме крови у борцов ниже, чем в контроле, и с повышением уровня мастерства от КМС до МС изменения не происходит.
Абсолютное количество орнитина в плазме крови у борцов ниже контроля, и с ростом мастерства до МС это количество снижается. Относительное количество орнитина так же ниже, чем в контроле, но при этом с ростом мастерства не меняется.
Абсолютное количество аргинина в плазме крови у борцов выше, чем в контроле, с ростом мастерства снижается. При этом относительное количество аргинина возрастает с ростом мастерства.
В общем, содержание глутамина и аргинина в полтора раза выше контроля, а у мастеров спорта содержание этих метаболитов практически нормализуется, даже становится ниже контроля. Так же снижено и продолжает снижаться количество орнитина. Скорее это связано с тем, что у мастеров спорта по борьбе становятся более мощными процессы детоксикации и обезвреживания аммиака. Как следствие уменьшается транспортная форма аммиака – глутамин и метаболит цикла синтеза мочевины – орнитин. Это же относится и относительным цифрам. А вот абсолютное количество аргинина у борцов выше контроля почти в полтора раза, а с ростом мастерства снижается почти до уровня контроля, при небольшом даже повышении относительного количества аргинина. Аргинин известен как нейропротектор, и нормализация с ростом мастерства говорит о снижении необходимости защищать нервную систему за счет усиления процессов детоксикации и обезвреживания аммиака в условиях реконструктивного протеолиза. Тем не менее организм борца относительное содержание аргинина сохраняет на высоком уровне и даже повышает его.
Таблица 2.29 Относительное содержание (%) аминокислот производных глутамата в крови у борцов в зависимости от квалификации (M+m(min-max)):
| Квалиф | Глутамин | Пролин | Орнитин | Аргинин |
| Контрольная | 15,8+0,9
(12,4-22,7) |
5,9+0,7
(3,0-10,8) |
3,1+0,6
(1,5-8,6) |
1,9+0,2
(0,8-2,9) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 19,1*+2,1
(14,9-22,7) |
3,8*+1,4
(1,0-5,5) |
2,3*+0,23
(1.5-3.1) |
2,1+0,3
(1,5-2,6) |
| МАСТЕРА СПОРТА | 13,5*+2,05
(4,10-20,07) |
4,4+2,1
(0,5-13,7) |
2,3*+0.22
(1,5-3,0) |
2,4*+0,3
(1,1-4,3) |
* Разница с контролем значима на уровне p<0.05.
** Разница с контролем значима на уровне p<0.01.
Общее относительное количество производных ГЛУ в крови у борцов уменьшается, за счет ГЛН.
Таблица 2.30 Абсолютное содержание (мкмоль/л (M+m(min-max)) разветвленных аминокислот в крови у борцов в зависимости от квалификации:
| Квалиф | Валин | Илейцин | Лейцин |
| Контрольная | 234,2+38,3
(106.3-458.5) |
43,9+5,3
(26.7-74.0) |
122,2+14,2
(78.9-206.6) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 202,4+7,3
(188.2-212.7) |
54,6+8,5
(37.8-64.5) |
132,8+10,1
(112,9-146,2) |
| МАСТЕРА СПОРТА | 172,3+47,8
(18.6-407.4) |
57,5+8,6
(28.0-94.3) |
130,1+15,4
(78,2-195,7) |
* Разница с контролем значима на уровне p<0.05.
** Разница с контролем значима на уровне p<0.01.
Абсолютное содержание разветвленных аминокислот у борцов представлено на таблице 2.30, относительное – на таблице 2.31.
Абсолютное и относительное количество разветвленных аминокислот у борцов не имеет достоверных отличий от контроля, с ростом мастерства достоверно не изменяется, за исключением роста относительного содержания изолейцина и лейцина.
Можно отметить рост изолейцина. Он стимулирует образование гемоглобина, что крайне важно для борцов. И еще, если общее абсолютное количество разветвленных аминокислот уменьшается, то относительное – увеличивается, то есть для организма борца важнее абсолютное количество этих метаболитов.
Таблица 2.31 Относительное содержание (%) разветвленных аминокислот в крови у борцов в зависимости от квалификации (M+m(min-max)):
| Квалиф | Валин | илейцин | Лейцин |
| Контрольная | 7,5+0,8
(3.8-1.9) |
1,5+0,2
(0.8-3.0) |
4,0+0,3
(2,7-6,0) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 5,1*+0,4
(4.3-5.6) |
1,3+0,2
(1,0-1,8) |
3,4*+0,4
(2,9-4,2) |
| МАСТЕРА СПОРТА | 6,6+1,8
(0.0-15.8) |
2,0*+0,2
(1,1-2,8) |
4,7*+0,4
(3,3-6,7) |
* Разница с контролем значима на уровне p<0.05.
** Разница с контролем значима на уровне p<0.01.
Содержание циклических аминокислот в плазме крови у борцов представлены на таблице 2. 32 (абсолютное) и 2.33 (относительное).
Абсолютное количество тирозина у борцов не отличается от контроля, при этом отмечается тенденция к снижению тирозина с ростом квалификации от КМС до МС. При рассмотрении относительного содержания тирозина в крови у борцов выясняется, что она с ростом квалификации имеет тенденцию к росту, что ясности не добавляет.
Таблица 2.32 Абсолютное (мкмоль/л (M+m(min-max)) содержание циклических аминокислот в крови у борцов в зависимости от квалификации:
| Квалиф | Тирозин | Фенилаланин | Триптофан | Гистидин |
| Контрольная | 55,2+5.5
(36.4-92.7) |
47,1+4.6
(21.2-78.9) |
30,1+8.4
(4.8-74.9) |
83,3+6.2
(56.0-111.7) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 61,4+3.7
(57.5-64.9) |
70,3*+10.0
(53.1-90.7) |
15,5*+4,5
(7.1-22.8) |
111,2*+5,0
(102.0-119.5) |
| МАСТЕРА СПОРТА | 52,0+4.6
(32.3-69.6) |
49,5+5,3
(19.3-64.9) |
2,7**+1.7
(0.4-11.0) |
84,3+12.7
(45.6-135.2) |
* Разница с контролем значима на уровне p<0.05.
** Разница с контролем значима на уровне p<0.01.
Содержание фенилаланина у борцов не отличимо от контроля, хотя у кандидатов в мастера абсолютное его количество выше контроля, а у мастеров – неотличимо от контроля, то есть снижается. При этом относительное содержание фенилаланина у борцов с ростом квалификации совершенно не меняется.
Из циклических аминокислот у борцов выделяется триптофан. Его количество у кандидатов в мастера спорта вдвое ниже, чем в контроле, а у мастеров спорта количество триптофана на порядок ниже, чем в контроле. Аналогична динамика относительного содержания триптофана – в целом ниже, чем в контроле, и у мастеров спорта ниже, чем у кандидатов в мастера.
Асолютное коилчество гистидина в крови у борцов с ростом квалификации снижается, относительное – не меняется.
Таблица 2.33 Относительное (%) содержание циклических аминокислот в крови у борцов в зависимости от квалификации(M+-m ( min- max)):
| Квалиф | Тирозин | фенилаланин | Триптофан | Гистидин |
| Контрольная | 1,8+0,1
(1.3-2.6) |
1,6+0,2
(0.8-3.0) |
0,8+0,2
(0.4-2.0) |
2,8+0,2
(2.0-4.9) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 1,5+0.3
(1.2-1.8) |
1,8+0.3
(1.1-2.4) |
0,4*+0.2
(0.1-0.6) |
2,8+0.6
(2.1-3.5) |
| МАСТЕРА СПОРТА | 1,8+0.3
(1.2-2.3) |
1,8+0.7
(1.1-3.5) |
0,2**+0.1
(0.0-0.4) |
3,0+0.8
(1.7-3.7) |
* Разница с контролем значима на уровне p<0.05.
** Разница с контролем значима на уровне p<0.01.
Суммарное количество аминокислот у борцов не имеет достоверных отличий от контроля. Но при сопоставлении этой величины у кандидатов и мастеров спорта обнаруживается, что у КМС суммарное количество аминокислот больше, чем в контроле, а с возрастанием мастерства достоверно снижается почти вдвое.
Абсолютное и относительное содержание серина, глицина и глутамина в плазме крови у борцов снижается.
Абсолютное содержание в плазме крови у борцов таурина, цистеина, орнитина, тирозина, фенилаланина, триптофана и гистидина снижается, при этом относительное их количество достоверно не изменяется. То есть метаболизм у борцов происходит таким образом, что снижение интенсивности реконструктивного протеолиза происходит при сохранении баланса именно этих аминокислот.
Абсолютное содержание в плазме крови у борцов глутаминовой кислоты, метионина, пролина, аргинина, валина, изолейцина и лейцина с возрастанием мастерства не изменяется. Относительное содержание в крови у борцов этих аминокислот так же не изменяется, за исключением лейцина и изолейцина, относительное количество, которых с возрастанием мастерства от КМС до МС даже возрастает.
И, наконец, единственная аминокислота, чьё количество возрастает и относительно и абсолютно – это аспарагиновая кислота. Ниже мы покажем, значение аспарагиновой кислоты для борцов действительно велико.
Хоккей
Изменений количества аланина у хоккеистов ни по абсолютным ни по относительным значениям в крови не отмечено поэтому его содержание в таблицах не приводится.
Таблица 2.34 Абсолютное (мкмоль/л (M+m(min-max)) содержание группы «древних» аминокислот в крови у хоккеистов в зависимости от квалификации:
| Квалиф | Аспарагиновая | Серин | Глутаминовая | Глицин |
| Контрольная | 58,5+14,3
(14.2-195.2) |
192,2+27,3
(64.0-358.0) |
135,0+29,5
(12.2-318.8) |
366,7+47,9
(126.9-586.1) |
| 1 разряд | 30,5*+11,1
(8,5-44,7) |
202,1+66,6
(133.5-335.5) |
137,6+18,3
(104.8-168.1) |
500,6*+82.1
(325.5-689.1) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 35,7+17,2
(6.3-103.2) |
186,4+33,3
(91.4-898.8) |
126,9+43,7
(31.1-289.3) |
428,6+98,8
(184.0-792.9) |
| МАСТЕРА СПОРТА | 46,1+25.6
(4.9-95.1) |
199,3+64,8
(71.2-280.3) |
117,8+12,7
(93.8-137.4) |
476,1+115,9
(231.2-652.1) |
* Разница с контролем значима на уровне p<0.05.
** Разница с контролем значима на уровне p<0.01.
Абсолютное и относительное содержание древних аминокислот в плазме крови у хоккеистов представлено в таблицах 2.34 (абсолютное) и 2.35 (относительное).
И абсолютное, и относительное количество аспарагиновой кислоты в крови хоккеистов в полтора раза ниже, чем в контроле. С ростом же квалификации от первого разряда до мастера спорта эти показатели растут, впрочем, так и не достигая уровня контроля.
И абсолютное и относительное количество серина в плазме крови у хоккеистов не отличается от контроля, с ростом квалификации не изменяется.
Абсолютное и относительное количество глутаминовой кислоты у хоккеистов не отличается от контроля, с возрастанием квалификации также не меняется.
Абсолютное содержание глицина в плазме крови у хоккеистов достоверно выше, чем в контроле. У перворазрядников уровень глицина выше, чем в контроле, у кандидатов в мастера и мастеров спорта уровень глицина не имеет достоверных отличий от контроля. Относительное содержание глицина у хоккеистов не отличается от контроля. У мастеров спорта относительное содержание глицина выше, чем в контроле, у перворазрядников и кандидатов в мастера не отличается от контроля.
Таблица 2.35 Относительное (%) содержание группы «древних» аминокислот в крови у хоккеистов в зависимости от квалификацииь (M+m(min-max)):
| Квалиф | Аспарагиновая | Серин | Глутаминовая | Глицин | Аланин |
| Контрольная | 1,7+0,3
(0.4-3.9) |
6,1+0,5
(3.7-8.2) |
3,9+0,5
(0.7-7.1) |
11,5+0,9
(7.9-15.7) |
12,5+1,1
(7,7-17,5) |
| 1 разряд | 0,95*+0,4
(,18-1,3) |
5,2+0,9
(3,9-7,0) |
3,8+0,5
(2,9-4,9) |
13,4+0,97
(11,49-14,44) |
13,5+0,6
(12,33-15,92) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 1,17+0,4
(0,23-3,3) |
5,9+0,15
(5,36-6,23) |
4,33+1,3
(1,06-9,23) |
13,1+1,06
(9,95-16,53) |
14,16+1,1
(11,89-18,4) |
| МАСТЕРА СПОРТА | 1,3+0,7
(0,43-2,75) |
6,5+0.7
(4,49-8,1) |
5,5+2,5
(2,09-10,5) |
14,21*+0,8
(9,95-20,03) |
11,8+1,3
(10-14,43) |
* Разница с контролем значима на уровне p<0.05.
** Разница с контролем значима на уровне p<0.01.
Таблица 2.36 Абсолютное содержание (мкмоль/л (M+m(min-max)) серосодержащих аминокислот в крови у хоккеистов в зависимости от квалификации:
| Квалиф | Таурин | Цистеин | Метионин |
| Контрольная | 60,7+13,6
(4.0-121.1) |
76,4+11,8
(7.6-131.9) |
25,5+2,3
(16.4-43.8) |
| 1 разряд | 78,9+13,6
(55.6-102.8) |
163,1**+15,3
(141.73-192.8) |
41,1*+6.7
(33.4-55.8) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 57,4+25,2
(14.0-137.2) |
113,7 *+24,9
(76.2-212.5) |
29,9+9,6
(6.2-62.3) |
| МАСТЕРА СПОРТА | 52,4+23,2
(17.0-96.2) |
115,1*+26.0
(48.2-197.9) |
34,0+13.1
(16.2-60.1) |
* Разница с контролем значима на уровне p< 0.05.
** Разница с контролем значима на уровне p<0.01.
Содержание серосодержащих аминокислот в плазме крови хоккеистов представлено на таблицах 2.36 (абсолютное) и 2.37 (относительное).
Абсолютное и относительное содержание таурина у хоккеистов не отличается от контроля, с ростом квалификации изменений относительно контроля нет.
Абсолютное содержание цистеина у хоккеистов в полтора раза превышает таковое в контроле. У перворазрядников содержание цистеина в крови выше контроля вдвое, у кандидатов и мастеров спорта абсолютное количество цистеина превышает таковое в контроле всего лишь в полтора раза. То же самое можно сказать и об относительном содержании цистеина.
Абсолютное содержание метионина у хоккеистов в полтора раза больше, чем в контроле. У перворазрядников – почти вдвое, а у кандидатов и мастеров спорта не отличается от контроля. Относительное же количество метионина у хоккеистов не отличается от контроля и с ростом квалификации не меняется.
Таблица 2.37 Относительное содержание (%) серосодержащих аминокислот в крови у хоккеистов в зависимости от квалификации (M+m(min-max)):
| Квалиф | Таурин | цистеин | Метионин |
| Контрольная | 1,8+0,3
(0.5-4.1) |
2,5+0,5
(0.8-3.5) |
0,9+0,2
(0.0-1.9) |
| 1 разряд | 2,30+0,66
(1,64-3,63) |
4,5*+0,45
(4,04-5,47) |
1,1+0,05
(1,01-1,18) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 1,6+0,56
(0,4-3,11) |
3,81+0,6
(2,51-6,12) |
0,9+0,2
(0,2-1,38) |
| МАСТЕРА СПОРТА | 1,7+0,53
(0,98-2,78) |
3,8+0,47
(2,87-4,41) |
1,1+0,2
(0,74-1,41) |
* Разница с контролем значима на уровне p <0.05.
** Разница с контролем значима на уровне p <0.01.
Таблица 2.38 Абсолютное содержание аминокислот производных глутаминовой кислоты в крови у хоккеистов (мкмоль/л (M+m(min-max)) в зависимости от квалификации:
| Квалиф | Глутамин | пролин | Орнитин | Аргинин |
| Контрольная | 498,1+61,0
(228.0-837.4) |
185,6+27,5
(52.3-330.7) |
99,6+21,4
(42.2-263.1) |
55,7+5,7
(25.0-77.0) |
| 1 разряд | 712,1*+87.9
(512.3-1066.6) |
47,1*+10,7
(28.3-65.4) |
111,4+8,33
(95.1-125.7) |
79,5*+6,71
(67.1-90.2) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 480,5+89,51
(235.6-759.8) |
162,3+47,2
(56.0-318,5) |
105,3+33.5
(31,7-244.6) |
56,4+10.5
(27.0-89.6) |
| МАСТЕРА СПОРТА | 541,8+139,8
(155.5-794.9) |
64,9*+32,8
(11.5-134.2) |
62,1+18,9
(24.7-85.2) |
67,7+38,8
(15.8-145.8) |
* Разница с контролем значима на уровне p <0.05.
** Разница с контролем значима на уровне p <0.01.
Содержание аминокислот, производных от глутаминовой в плазме крови хоккеистов представлено в таблице №38 (абсолютное) и таблице №39 (относительное).
Абсолютное и относительное содержание глутамина в крови у хоккеистов не отличается от контроля. Тем не менее, у перворазрядников содержание глутамина (абсолюное и относительное) достоверно выше, чем в контроле.
Абсолютное и относительное содержание пролина в плазме крови у хоккеистов вдвое ниже, чем в контроле. У перворазрядников и мастеров спорта абсолютное содержание пролина втрое ниже, чем в контроле, а у кандидатов в мастера – не отличается от контроля.
Абсолютное и относительное содержание орнитина в плазме крови у хоккеистов неотличимо от контроля. Представители всех квалификационных групп так же имеют неотличимое от контроля абсолютное содержание орнитина. Исключение составляют мастера спорта. У них относительное количество орнитина ниже, чем в контроле.
Абсолютное содержание аргинина в плазме крови у хоккеистов не отличается от контроля. У перворазрядников содержание аргинина повышено, по сравнению с контролем. Относительное содержание аргинина в любой квалификационной группе не отличается от такового в контроле.
Таблица 2.39 Относительное содержание (%) производных глутаминовой кислоты в крови хоккеистов в зависимости от квалификации (M+m(min-max)):
| Квалиф | Глутамин | пролин | орнитин | Аргинин |
| Контрольная | 15,8+0,9
(12.4-22.7) |
5,9+0,7
(3.0-10.8) |
3,1+0,6
(1.5-8.6) |
1,9+0,2
(0.8-2.9) |
| 1 разряд | 19,04*+2,11
(15,11-22,35) |
1,31+0.31
(1,0-1,93) |
3,14+0.35
(2,44-3,62) |
2,2+0.31
(1,89-2,87) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 15,27+0.47
(13,48-16,6) |
5,08+1,11
(2,64-9,1) |
3,02+0,56
(2,08-5,10) |
1,83+0,2
(1,32-2,56) |
| МАСТЕРА СПОРТА | 16,9+1,78
(13,48-19,5) |
1,8+0,6
(1,0-2,99) |
2,08*+0,09
(1,9-2,21) |
1,94+0,65
(1,2-3,25) |
* Разница с контролем значима на уровне p <0.05.
** Разница с контролем значима на уровне p <0.01.
Таблица 2.40 Абсолютное содержание разветвленных аминокислот в крови у хоккеистов в (мкмоль/л (M+m (min-max)) в зависимости от квалификации:
| Квалиф | Валин | Изолейцин | Лейцин |
| Контрольная | 234,2+38,3
(106.3-458.5) |
43,9+6,3
(26.7-74.0) |
122,2+14,2
(78.9-206.6) |
| 1 разряд | 181,8+32.4
(159.5-219.0) |
69,1*+8.7
(54.1-90.2) |
145,6+25.0
(96.6-185.1) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 191,2+67.5
(91.1-243.3) |
57,0+38.5
(20.0-101.2) |
120,7+34.4
(72.4-169.3) |
| МАСТЕРА СПОРТА | 149,5*+54.1
(70.8-218.1) |
41,7+21.3
(10.5-73.1) |
99,0+32.8
(28.1-173.7) |
* Разница с контролем значима на уровне p<0.05.
** Разница с контролем значима на уровне p<0.01.
Содержание разветвленных аминокислот в плазме крови хоккеистов представлено в таблице 2.40 (абсолютное) и 2.41 (относительное).
В крови у хоккеистов абсолютное количество валина ниже, чем в контроле. При этом у перворазрядников и кандидатов в мастера спорта абсолютное содержание валина неотличимо от контроля, а у мастеров спорта – ниже на треть.
Содержание лейцина и изолейцина, как абсолютное, так и относительное у хоккеистов не отличается от такового в контроле, за исключением перворазрядников, у которых абсолютное содержание изолейцина в полтора раза выше, чем в контроле.
Таблица 2.41 Относительное содержание (%) разветвленных аминокислот в крови у хоккеистов в зависимости от квалификации (M+m(min-max)):
| Квалиф | Валин | Илейцин | лейцин |
| Контрольная | 7,5+0,8
(3.8-11.9) |
1,5+0,2
(0.7-3.1) |
4,0+0,3
(2.7-6.0) |
| 1 разряд | 5,1*+1.1
(3.5-6.4) |
1,8+0.2
(1.8-1.9) |
3,9+0.5
(3.4-4.5) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 6,3+1.7
(4.8-9.1) |
1,8+1.0
(0.6-3.4) |
4,0+0.6
(3.2-4.7) |
| МАСТЕРА СПОРТА | 5,2*+0.8
(4.6-6.1) |
1,2+0.3
(0.9-1.6) |
3,0+0.7
(2.4-3.8) |
* Разница с контролем значима на уровне p<0.05.
** Разница с контролем значима на уровне p <0.01.
Таблица 2.42 Абсолютное содержание циклических аминокислот в крови у хоккеистов в зависимости от квалификации (мкмоль/л(M+m(min-max)):
| Квалиф | Тирозин | Фенилаланин | Триптофан | Гистидин |
| Контрольная | 55,2+5,5
(36.4-92.7) |
47,1+4,6
(21.2-78.9) |
30,1+8,4
(4.8-74.9) |
83,3+6,2
(56.0-111.7) |
| 1 разряд | 80,4*+16.9
(60.6-111.2) |
82,6*+25.1
(48.7-118.8) |
17,4*+2.6
(10.5-23.8) |
125,9*+15.7
(109.5-155.5) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 62,8+25.4
(41.7-103.6) |
50,3+14.5
(40.5-15.7) |
13,4**+2.9
(2,8-26.6) |
75,0+18.3
(48.9-119.4) |
| МАСТЕРА СПОРТА | 59,4+28.3
(37.8-91.5) |
52,8+13.2
(31.5-77.6) |
7,2**+4.4
(0.9-18.5) |
94,6+36.5
(35.0-166.5) |
* Разница с контролем значима на уровне p <0.05.
** Разница с контролем значима на уровне p <0.01.
Содержание циклических аминокислот в плазме крови у хоккеистов представлено в таблице 2.42 (абсолютное) и 2.43 (относительное).
Абсолютное содержание тирозина у хоккеистов снижается с ростом квалификации. Относительное содержание тирозина с ростом квалификации у хоккеистов не изменяется.
Абсолютное содержание фенилаланина у хоккеистов снижается с ростом квалификации. Относительное содержание фенилаланина с ростом квалификации у хоккеистов не изменяется.
Абсолютное содержание триптофана у хоккеистов снижается с ростом квалификации. Относительное содержание триптофана с ростом квалификации у хоккеистов монотонно уменьшается.
Абсолютное содержание гистидина у хоккеистов снижается с ростом квалификации. Относительное содержание гистидина с ростом квалификации у хоккеистов не изменяется.
Таблица 2.43 Относительное содержание циклических аминокислот в крови у хоккеистов (%) в зависимости от квалификации (M+m(min-max)):
| Квалиф | Тирозин | фенилаланин | триптофан | Гистидин |
| Контрольная | 1,8+0,1
(1,3-2,6) |
1,6+0,2
(0,8-3,0) |
0,8+0,2
(0,4-2,0) |
2,8+-0,1
(2,0-4,9) |
| 1 разряд | 2,1+0.6
(1.7-2.4) |
2,1+0.4
(1.7-2.4) |
0,4*+0.1
(0.3-0.6) |
3,4+0.4
(3.2-3.9) |
| КАНДИДАТЫ В МАСТЕРА СПОРТА | 2,1+0.6
(1.3-2.7) |
1,7+0.5
(1.3-2.6) |
0,3**+0.1
(0.0-0.2) |
2,4+0.4
(1.9-3.2) |
| МАСТЕРА СПОРТА | 2,2+0.9
(1.4-3.2) |
1,9+0.6
(1.4-2.7) |
0,1**+0.2
(0.0-0.4) |
3,0+0.6
(2.3-3.7) |
* Разница с контролем значима на уровне p <0.05.
** Разница с контролем значима на уровне p <0.01.
Выше мы установили, что суммарное количество аминокислот в плазме крови хоккеистов минимально, по сравнению с другими исследованными группами. Баланс аминокислот, характерный для хоккеистов, определяется пониженным абсолютным, а, главное, относительным содержанием аспарагиновой кислоты, пролина, валина и триптофана, и повышенным содержанием цистеина.
У хоккеистов и среднее количество аминокислот в плазме крови и эта величина у представителей разного уровня квалификации не имеет достоверных отличий от контроля, за исключением перворазрядников, у которых суммарное количество аминокислот выше, чем в контроле (p<0,05). Тенденция к снижению количества аминокислот с ростом квалификации у хоккеистов вполне отчетливо видна.
Единственная аминокислота, чье количество и абсолютное, и относительное возрастает с ростом квалификации у хоккеистов — это аспарагиновая кислота.
Снижается с ростом квалификации содержание цистеина, орнитина и триптофана, тоже и абсолютное, и относительное.
Не изменяется с ростом квалификации в плазме крови у хоккеистов ни относительное ни абсолютное количество серина, таурина, метионина, пролина.
Содержание остальных аминокислот с ростом квалификации у хоккеистов абсолютное – снижается, а относительное – сохраняется на одном уровне. Это глутамин, аргинин, валин, изолейцин, лейцин, тирозин и фенилаланин.
Единственная аминокислота, чьё абсолютное содержание сохраняется на одном уровне, что на фоне снижения суммарного количества аминокислот приводит к возрастанию относительного количества – это глутаминовая кислота.
РЕЗЮМЕ ПО 2 ГЛАВЕ:
Таким образом, мы видим, что разница в содержаниии амнокислот и соотвеветственно в их метаболизме у представителей рассмотренных видов спорта и по мере роста мастерства проявляет себя в следующих особенностях.
Легкоатлеты:
У легкоатлетов чем выше мастерство, тем меньше аспарагиновой кислоты (у МСМК в 8 раз меньше, чем у КМС). По глутаминовой кислоте – обратная картина- у МСМК – в 2 больше чем у 1р, и в 1,5 у КМС.
Сумма аминокислот плазмы крови у МСМК в 2 раза меньше, чем у перворазрядников и КМС. Эти результаты согласуются с имеющимися данными литературы[197,260], что общее содержание аминокислот подвержено правилу повышения экономичности метаболизма по мере роста квалификации.
Соотношение аминокислот в большей степени отражает регуляторную сторону их обмена. Показатели относительного содержания аспарагиновой кислоты соответвтвуют таковым по абсолютным показателям, то же можно сказать о серине. Глутаминовая кислота дает тоже обратную картину. По глицину существенной разницы не выявлено. Сумма относительного количества древних аминокислот с ростом мастерствва достоверно не меняется. В то же время следует отметить, что по мере роста мастерства у легкоатлетов происходят изменения соотношения аспартата и глутамата в крови – снижение аспарагиновой и рост глутаминовой кислоты.
Серосодержащие аминокислоты
Таурин резко снижается у МСМК в 4-5 раз по отношению к другим группам, снижаетсся количество цистеина в 2 раза. Метионин хотя и имеет тенденцию к снижению, но незначительно. Снижение суммы абсолютного количества серосодержащих аминокислот с ростом мастерства отражает общую закономерность снижения интенсивности метаболизма в покое с ростом мастерства.
Относительное количество отражает аналогичную динамику абсолютного количества серосодержащих аминокислот с ростом мастерства. Особенно это относится к таурину.
Глутаминовая кислота и ее производные
Абсолютное содержание самой ГЛУ с ростом квалификации имеет тенденцию к повышению, а все ее производные – резко снижаются и по отдельности и по сумме. У МСМК абсолютное количество глутамина снижено в полнора раза, пролина в 7 раз орнитина в 5 раз, аргиниа в 4 раза но при этом содержание самой глутаминовой кислоты возрастает в 1,5 -2 раза по отношению к контролю. Глутамат образуется в мозге и мышцах, поэтому закономерно ее увеличение по мере проста мышечной массы у более тренированных спортсменов. Это же подтверждает увеличение относительного его количества в процентах. А по отдельным производным глутаминовой кислоты можно сказать, что несмотря на уменьшение общего количества глутамина в крови у МСМК относительное его количество увеличивается примерно в 1, 5 раза. Содержание остальных аминокислот производных глутамата снижается, как их относительное так и абсолютные значения. Что очевидно связано с особенностями изменений метаболизма и регуляторного значения в нем данных аминокислот.
Разветвленные аминокислоты
Наблюдается снижение всех по абсолютной величине, и остаются на одном уровне по относительной. Обращает на себя внимание, что количество валина у всех групп превосходит таковое у лиц, не занимающихся профессионально спортом. Валин можно рассматривать как универсальный метаболический ресурс необходитмый для повышения адаптивных возможностей легкоатлетов по мере роста их квалификации. В то время как количество лейцина и изолейцина по абсорлютным значениям по мере роста квалификации не изменяется а по относительным значениям уменьшается в полтора раза. То есть они не имеют такого значения для легкоатлетов в регулятоных механизмах как валин, но очевидно активно используются в анаболических процессах.
Циклические аминокислоты
Абсолютное значение по тирозину и фенилаланину в группе КМС и 1р. имеет тенденцию к росту в 1,5-2 раза, тогда как у МСМК примерно соответствует уровню контроля. На стадии становления необходим ресурс в крови метаболитов предшественников адреналовых и тиреооидных гормонов- гормонов стресса. А на уровне высокого мастерства они в таком количестве не требуются и их уровень снижается, особенно по отношению к КМС. То есть меняется метаболический профиль спорстмена по мере становления его как профессионала высокого уровня.
Динамика изменения количества триптофана и гистидина также имеет место. Первоначально происходит их существенное увеличение – в 3-4 раза.С ростом мастерства их количество в крови уменьшается. По триптофану разница между МСМК и всеми предшествующими группами; к контолю более чем на порядок, а по отношению к 1р., КМС и МС еще более выражена.
Триптофан – предшественник серотонина и кинуренина. Метаболизирование триптофана по кинурениновому пути необходимо для образования нуклеотидов, участвующих в образовании энергии (НАД и ФАД), по серотониновому- для образования серотонина – антистрессового метаболита и медиатора ЦНС. У МСМК высокоэффективны регуляторные процессы и видимо нет надобности в повышенных количествах означенных метаболитов. Или они активно используются и их расход повышен.
Гистидин – предшественник гистамина. Его количество возрастает в плазме крови перворазрядников в 2 раза, а затем по мере роста квалификации ступенчато снижается. У МСМК его уровень в крови статистически значимо не отличается от контроля
В целом следует отметить, что сумма циклических аминокислот резко снижена только у МСМК причем падение – резкое. Большое значение очевидно в данном случае имеет не сумма, а особенности метаболизма этих аминокислот по отдельности (Рис 4).
Рис 4. Преобладание аминокислот-адаптогенов в крови легкоатлетов различного уровня мастерства:
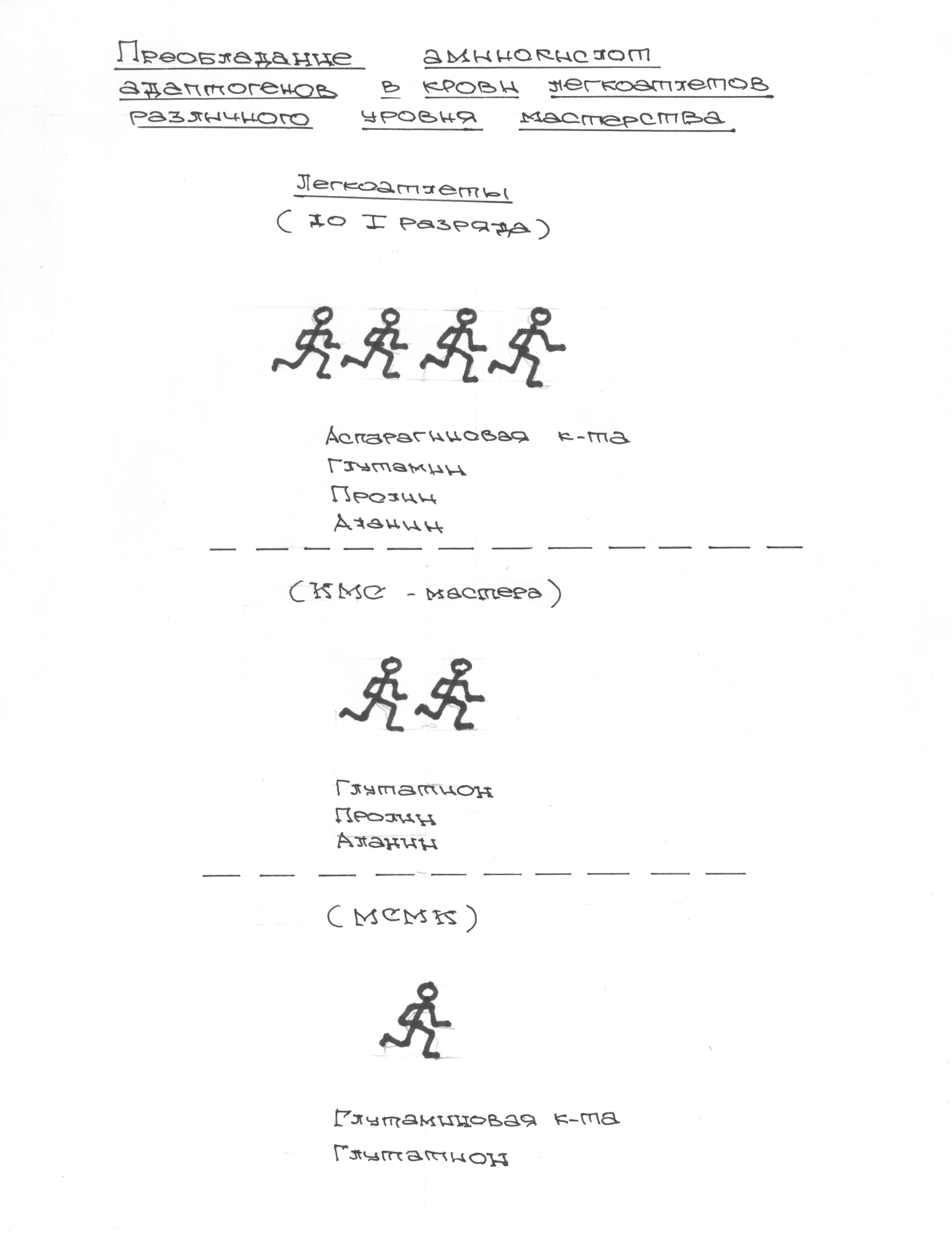
Борцы:
Обращает на себя внимание изменения количества дикарбоновых аминокислот по мере роста мастерства борцов. Количество аспарагиновой кисолты уменьшается у КМС по сравнению с контролем в 3 раза, а затем у МС оно снижается и примерно соответствует уровню контроля. Но при этом количество глутаминовой кислоты в 2 раза возрастает у КМС и остается примерно на том же уровне у МС.
Глутаминовая кислота является нейромедиатором, анаболиком и антиоксидантом. В большом количечтве образуется в мозге и мышцах. Судя по динамики ее количества у борцов глутамат является важнейшей аминокислотой в повышении адаптивных возможностей организма борца. Производное глутаминовой кислоты глутамин претерпевает похожую динамику. Его количество в крови борцов сначала растет, а затем приближается к уровню контроля. Увеличивается содержание такого производного глутамата как аргинин как у КМС, так и МС. Глутамин и аргинин является мощными адаптогенами и аанаболиками, что чрезвычайно важно для становления борца профессионала.
Возможно все это свзяно с более мощными процессами детоксикации, так как борцы часто испытывают гипоксии, что повышает значение обмена глутаминовой кислоты и ее производных.
Аспарагиновая кислота в больших количествах обнаружена в ЦНС, принимает участие в метаболизме азота, что для борца важно.
Серосодержащие и разветвленные аминокислоты существенных изменений их содержания по мере повышения мастерства борцов не претерпевают.
Циклические аминокислоты:
Отмечается снижение абсолютного количества у КМС фенилаланина в 2 раза к контролю и триптофана в 2 раза. Затем у МС количество фенилаланина нормализуется, а триптофана уменьшается еще в 7 раз. В относительных цифрах тоже снижение. То есть фенилаланин и триптофан так же как и у легкоатлетов имеет важное значение в росте уровня квалификации (Рис 5).
Рис. 5. Преобладание аминокислот-адаптогенов в крови борцов различного уровня мастерства:
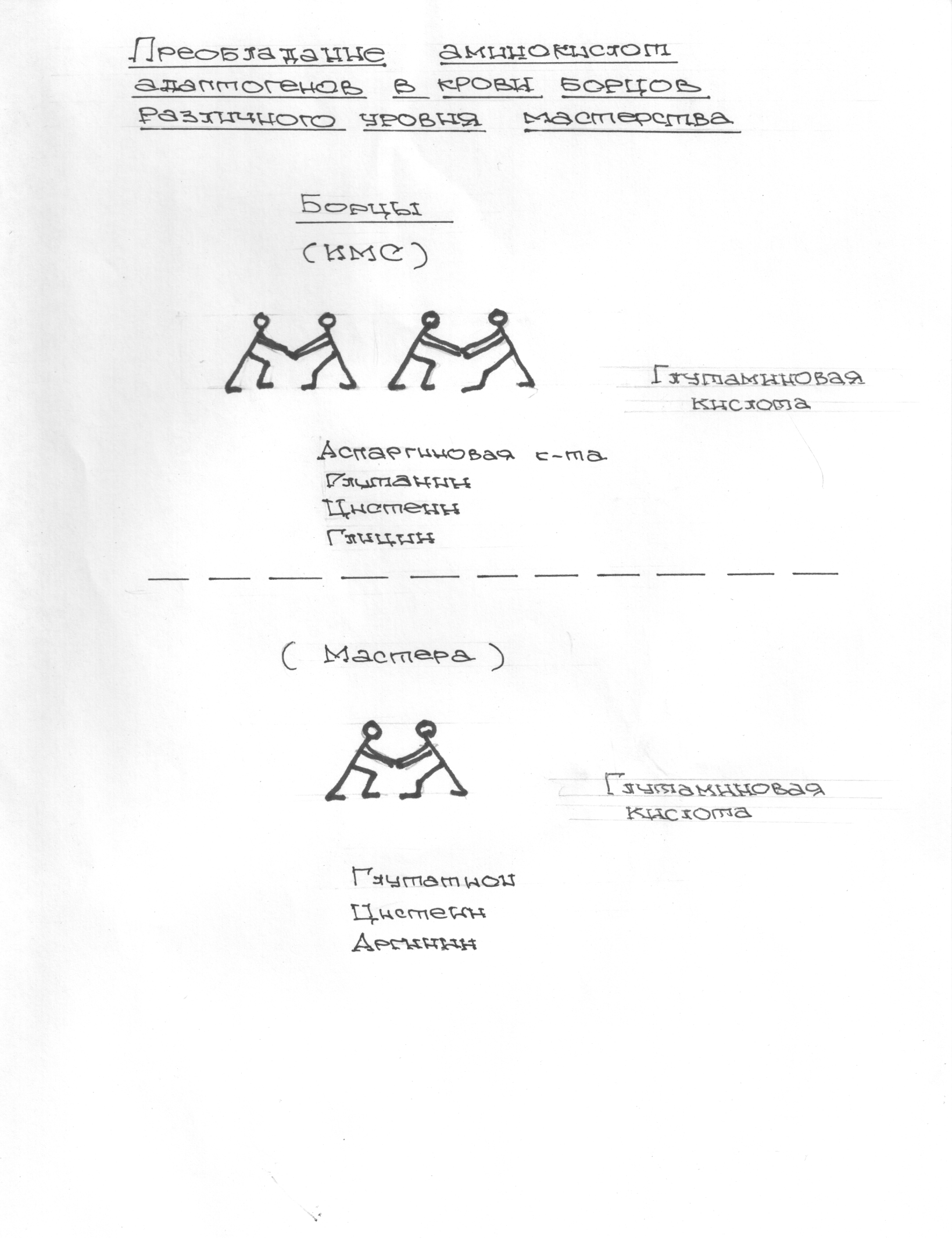
Хоккей:
Динамики содержания как по асолютным, так и по относителям показателям «древних» аминокислот нет по сравнению с контролем. Никакой достоверной разницы нет и между КМС и МС. За исключением аспартата – увеличение количества с ростом мастерства на треть только по абсолютным показателям. Отмечается так же повышение количества аргинина у начинающих спортсменов (1р.) которое затем примерно соответствует уровню контроля у КМС и МС.
С ростом мастерства абсолютное количество серосодержащих аимнокислот возрастает почти полтора раза по каждой аминокислоте, так и по сумме. То же и по относительным цифрам. Сначала отмечается тенденция к приросту (1р.), а затем наблюдается небольшое снижение по количеству таурина, который является универсальным тормозным медиатором и видимо препятствует росту мастерства в хоккее, очевидно, чтобы повысилась подвижность хоккеиста.
Примерно 2 раза как по абсолютным, так и по относительным показвателям отмечается прирост количества цистеина. Хоккеисты — аэробы, а это требует мощных антиоксидантных систем, которые зависят от образования глутатиона, зависимого в свою очередь от включения в них цистеина. А вот по метионину существенной разницы нет. Ресурс серы ненсмотря на активное ее включение в цистеин (и антиоксидантные механизмы) в организме сохраняется.
Группа глутамата. Количество самой глутаминовой кислоты с ростом мастерста существенно не изменяется, но при этом снижается количество глутамина в 1,5, что видимо связано с усилением анаболических процессов и детоксикации у более квалифицированных спортсменов. Растет количество пролина, что скорее всего связано с особенностями соединительной ткани.
Существенных изменений количества разветвленных аминокислот не отмечается.
Циклические аминокислоты.
Количество фенилаланина и тирозина у перворазрядников возрастает в 1,5-2 раза к контролю, а затем снижается примерно на треть и у КМС и МС от контроля не отличается. Что было характерно и для легкоатлетов, и для борцов.
Количество триптофана снижается у 1р. примерно в 2 раза к контролюкак как по абсолютным, так и по относительным показателям, а затем у КМС и МС снижается еще больше. Количество гистидина сначала приростает примерно в 1,5 раза, а затем нормализуется. То есть примерно так же как у предыдущих групп спортсменов (Рис 6).
Рис. 6. Преобладание аминокислот-адаптогенов в крови хоккеистов различного уровня мастерства:

Таким образом, Каждый вид спорта имеет свой определенный метаболический профиль с преобладанием определенной ааминокислоты или группы аминокислот. Суммировать полученные результаты можно в следующих выводах:
-
В процессе становления спортсмена-профессионала в совершенствовании его адаптивных систем в разных видах спорта участвуют различные аминокислоты-адаптогены.
-
Суммарное количество аминокислот обратнопропорционально интенсивности стрессовой реакции на спортивную физическую нагрузку.
-
Наиболее длительно повышенный стрессовй фон( судя по повышенному уровню суммы аминокислот в плазме крови) наблюдается у легкоатлетов, до уровня МСМК.
-
Содержание абсолютного и относительного количества отдельных аминокислот может иметь различное значение в совершенствовании адаптивных механизмов к спортивным нагрузкам, характерным для каждого вида спорта по мере возрастания квалификации.
ГЛАВА З. МЕХАНИЗМЫ ВЛИЯНИЯ СВОБОДНЫХ АМИНОКИСЛОТ НА РАЗЛИЧНЫЕ УРОВНИ АДАПТИВНЫХ МЕХАНИЗМОВ ОРГАНИЗМА СПОРТСМЕНА НА ОСНОВАНИИ КОРРЕЛЯЦИОННОГО АНАЛИЗА
ВВЕДЕНИЕ
Исходя из полученных результатов все группы исследованных спортсменов имеют свои особенности по содержанию и соотношению исследованных аминокислот – аминокислотный профиль, который может влиять на морфологию, на физиологию и на психологию. Причем этот профиль меняется с ростом мастерства спортсмена, но не выходит за рамки вида спорта. Аминокислоты лежат в основе регуляторных возможностей организма. Что позволяет делать прогноз в соответствии с количеством свободных аминокислот и изменением их количества в процессе тренировок. Применение аминокислот должно производиться на основе знания метаболического профиля характерного для данного вида спорта.
Представители любого вида спорта являются в значительной степени мезоморфами (классификация Шелдона). Они ведут одинаковый образ жизни, занимаются деятельностью одного рода, достигли определенного и сравнимого уровня мастерства. Разными являются характер физической нагрузки и характер деятельности. Легкоатлеты ведут борьбу главным образом с несовершенством собственного тела, борцы противостоят противнику в непосредственном единоборстве, хоккеисты действуют в составе команды. Вот эта разница в характере деятельности и связанным с ней характером физической нагрузки видимо и приводят к тому, что аминокислотный состав плазмы крови имеет определенные, характерные для данного вида спрта значения
Формирование тела изначально базируется на наследственных свойствах организма, но по мере развития все большее значение обретают приобретенные в онтогенезе качества. Индивидуальные особенности морфологических, физиологических, биохимических, психологических параметров человека исследованы достаточно широко. Овладение опорно-двигательным аппаратом происходит через формирование двигательных навыков. Каждый двигательный навык на каждом уровне по Бернштейну имеет как минимум два компонента – собственно двигательный и вегетативный. Соответственно, двигательные стереотипы так же состоят из двух компонентов. Это физиологический уровень. Сформированный двигательный стереотип сопровождается вегетативным компонентом, сформированным для обеспечения движения. При этом на уровне столкновения интересов целого органа и отдельных его структурно-функциональных единиц формируется биохимический, метаболический стереотип. Мы считаем, что можно говорить по крайней мере о двух стереотипах: аэробный-анаэробный и аминокислотный. Первый имеет отношение к энергообеспечению, второй – к возбудимости клетки, регуляции ее метаболизма. Соотношение этих метаболических стереотипов и определяет, в конечном итоге суммарный метаболический стереотип.
Аминокислотный метаболический стереотип первичен по отношению к энергетическому. Дело в том, что энергетический стереотип формируется в соответствии с характером физической нагрузки, каковой, в свою очередь, определяется образом жизни, а аминокислотный стереотип связан с качествами нервной системы (а даже наоборот, аминокислотный метаболический стереотип определяет качества нервной системы), которые принято считать наследственными. Ранее мы отмечали, что на основании результатов исследований на животных и больных в клинике можно предположительно выделить «МОЗГОВЫЕ» и «ПЕЧЕНОЧНЫЕ» адаптивные типы. В первом случае в плазме крови и других тканях организма пробладают аминокислоты и их производные образующиеся в печени, во-вторм- в ткани морзга[237].
С другой стороны, мы имеем данные о том, что аминокислотный метаболический стереотип тоже изменяется под влиянием физической нагрузки. Нами установлена связь между относительной мышечной массой и относительным содержанием в крови глутаминовой кислоты. Характер кривой зависимости одинаков у представителей разных видов спорта, но различается по уровню содержания глутаминовой кислоты. 49% мышечной массы является порогом для резкого повышения содержания глутаминовой кислоты в крови. Мышечная масса зависима от характера физической нагрузки, следовательно, и содержание глутаминовой кислоты зависит от характера физической нагрузки.
Энергообеспечение зависит от характера физической нагрузки.
Все эти стереотипы – двигательный, с его двигательным и вегетативным компонентами, метаболический энергетический и метаболический аминокислотный формируют морфологическую структуру тела, что отражается в компонентном составе тела. Компонентный состав тела, таким образом, является производным от совокупного метаболического стереотипа с одной стороны, и, опосредованно, от характера физической нагрузки и, в меньшей степени, от характера движений.
Классические исследования Кречмера, Шелдона и других прямо показывают, что разные аспекты человеческой сущности имеют связь. Классификации конституции включают в себя как морфологические, так и физиологические признаки
В классических исследованиях Мерлина, что характер движений зависит от вида деятельности и все в совокупности формирует индивидуальный стиль деятельности.
Психологи утверждают, что индивидуальный стиль деятельности определяется темпераментом, типом нервной системы.
И.П. Павлов помимо темперамента выделял еще три «чисто человеческих типа» высшей нервной деятельности: мыслительный, художественный, средний.
Мыслительный (преобладает активность второй сигнальной системы мозга левого полушария) — рассудительны, склонны к детальному анализу явлений, к отвлеченному абстрактно-логическому мышлению. Умеренны в своих чувствах, сдержанны, обычно интересуются математикой, философией, склонны к научной деятельности.
Художественный тип (преобладает активность первой сигнальной системы мозга правого полушария) — мышление образное, эмоциональное живое восприятие действительности, стремятся к широкому кругу общения, типичные лирики, людей мыслительного типа рассматривают как «сухарей». Интересуются театром, искусством, поэзией, музыкой.
Средний тип (80%) – равновесное сочетание рационального и эмоционального, одно над другим может незначительно преобладать. По мнению И.П. Павлова преобладание типа высшей нервной деятельности зависит от воспитания в детстве (до 12-16 лет).
Голубева Э.А. (1980, 2000) исследуя механизмы памяти, говорит о влиянии свойств нервной системы на механизмы запоминания. Речь идет о таких свойствах, как сила-слабость, инертность-лабильность и уравновешенность.
Наибольший интерес представляет собой классификация Шелдона. В своей работе он установил связь между эмбриогенезом, морфологическими признаками тела и психологическими характеристиками.
У Шелдона в основе классификации лежит преобладание в развитии того или иного зародышевого листка. В соответствии с этим он выделяет эндоморфию, мезоморфию и эктоморфию как преобладание зародышевых листков в строении тела.
Эндоморфия выражается в висцеротонии, мезоморфия – в соматотонии, и эктоморфия – в церебротонии. Классификация Шелдона хорошо согласуется с классификацией Кречмера.
Таким образом, стремление установить границы изменчивости в пределах вида «человек» издавна наблюдалось в научном мире. Многочисленные исследования влияния наследственности на индивидуальность дополняют картину.
Тем не менее, вопрос о соотношении между метаболическими, физиологическими, морфологическими и психологическими параметрами индивида оставался открытым.
Трудности были связаны, прежде всего, с трудностью создания групп, однородных по полу и возрасту, все представители которых ведут одинаковый образ жизни, но имеют различие, ну скажем, по характеру нагрузки.
Вторая трудность – оценить психологические последствия для личности соответствия или несоответствия наследственных задатков организма для данной деятельности. Морфологические, физиологические или биохимические параметры организма в какой-то степени меняются, подстраивая организм под деятельность.
Формирование тела человека происходит в соответствии с наследственной программой, записанной в хромосомах. Влияние среды вносит коррекции в реализацию наследственной программы. Состояние внутренней среды организма согласуется с состоянием внешней среды при посредстве опорно-двигательного аппарата с одной стороны, и метаболизма – с другой стороны.
Исходя из полученных нами результатов в формировании всех уровней апдаптивных механизмов человека принимают участие все те же самые древнейшие прародители и регуляторы живой материи – аминокислоты. Они как бы создают некие стержни, на которые насаживаются адаптивные системы начиная от морфологического строения организма, затем его физиологические возможности и наконец особеннности его психической деятельности.
Мы выяснили, что характер деятельности представителей разных видов спорта определенным образом связан с особенностями метаболизма, выраженных в различии абсолютного и относительного содержания аминокислот – с одной стороны, и разно направленной динамики содержания аминокислот по мере роста квалификации, с другой.
Суммарное количество аминокислот есть субстратная основа для формирования структурного следа адаптации. Структурный след адаптации формируется у представителей разных видов спорта для обеспечения деятельности, характерной для вида спорта. Этот след остается и в строении тела, выявляется при функциональной диагностике состояния внутренних органов, и при тестировании психологических характеристик, что хорошо известно каждому тренеру.
Выявленное различие в метаболизме аминокислот поставило перед нами задачу – выяснить связи аминокислот с формированием конкретных признаков структурного следа адаптации.
Мы провели морфометрическое, физиологическое и психологическое исследование у легкоатлетов, борцов и хоккеистов, и исследовали корреляционные связи между содержанием аминокислот в плазме крови. Исследованию морфометрических, физиологических особенностей кардиореспираторной системы, психологических особенностей у спортсменов в данных видах спорта посвящено огромное количество работ. Поэтому мы не ставили своей задачей сравнивать эти показатели и их изменения по мере роста спортивного мастерства. А вот роль амнокислот в регуляции этих уровней адаптивных механизмов спортсмена исследована слабо, хотя они широко, порою неоправданно, применяются в спорте и в фитнесе. Поэтому мы попытались выявить закономерности регуляторного воздействия аминокислот на рассматриваемые уровни адаптивных механизмов.
Исследование проводилось комплексно. Спортсмены приходили на исследование утром, проводилось психологическое тестирование, затем морфологическое исследование, определение параметров гемодинамики в покое (исследование проводилось на протяжении пятисот ударов сердца), далее проводился нагрузочный тест со ступенчатым повышением нагрузки, при этом исследовались функции дыхательной и сердечно-сосудистой системы.
3.1 Корреляции показателей морфометрии и содержания свободных аминокислот плазмы крови у представителей разных видов спорта
Морфологическое исследование мы проводили по методике Матейко. Были определены следующие параметры: рост, вес, длина конечностей (длина рук (ДР), длина ног (ДН), длина туловища (ДТ), ширина плеч (ШП), ширина таза (ШТ). Были вычислены: индекс пропорциональности (ИП), весоростовой индекс (ВР), относительная длина конечностей (относительная длина рук (ДРО), относительная длина ног (ДНО), абсолютные и относительные веса жирового (ЖК, ЖКО), мышечного (МК, МКО), и костного (КК, ККО) компонентов тела. Результаты морфометрического исследования представлены в ПРИЛОЖЕНИИ. Так как динамика морфометрических показатьелей в рассматриваемых видах спорта хорошо исследована, то мы не ставили перед собой задачу сопоставления полученных результатов, а сосредаточились на исследовании корреляционных связей морфометрических показателей и динамики свободных аминокислот в плазме крови спортсменов.
Корреляционные связи были посчитаны с помощью программы SPSS, учитывались только сильные достоверные корреляционые связи с достоверностью не ниже 0,05. Результаты статистического исследования представлены в ПРИЛОЖЕНИИ.
3.1.1 Корреляции морфометрических показателей с содержанием аминокислот у борцов
В материале, представленном ниже, мы демонстрируем корреляционные связи у борцов между абсолютным (таблица 3.1) и относительным (таблица 3.2) количеством аминокислот и морфометрическими показателями.
Таблица 3.1 корреляционные связи абсолютного содержания аминокислот и морфологических показателей у борцов:
| Показатели | «Древние» аминокислоты | Серосодержащие аминокислоты | Произв. ГЛУ | Разветвленные
аминокислоты |
Циклические
аминокислоты |
| Возраст | АСП, -ГЛИ | ||||
| Рост | АСП | ||||
| ВР | АСП | ||||
| ИП | АЛА | -ОРН | |||
| ШП | -ОРН | ||||
| ШПО | -ТАУ | ||||
| ШТ | АСП | ПРО | |||
| ШТО | АРГ | ||||
| ДР | АСП | ОРН | |||
| ДРО | -ОРН | -ТРИ | |||
| ДН | АСП | ПРО | |||
| ЖК | АСП,-АЛА | ||||
| ЖКО | АСП | ||||
| МК | АСП | ||||
| МКО | ГЛУ | ||||
| ККО | -АСП, ГЛИ | ||||
| ИТОГО 24 | 15 | 1 | 7 | 1 |
На таблице 3.1 обращает на себя внимание то, что рекордсменами по количеству корреляций с морфологическими показателями у борцов являются древние аминокислоты- всего 15 достоверных сильных корреляций из общего количества 24. Среди древних аминокислот наибольшее количество корреляций дает аспарагиновая кислота -10 корреляций. На втором месте – производные глутаминовой кислоты. Орнитин, пролин и аргинин – 4, 2 и 1 корреляции соответственно. Кроме того, по одной связи отмечено у серосодержащих (таурин) и циклических (триптофан) аминокислот.
При этом аспарагиновая кислота имеет 6 корреляций с абсолютными размерами тела борца, одну связь с возрастом, все корреляции положительны, а также 3 корреляции с относительными показателями. Иначе говоря, аспарагиновая кислота оказывает значительное влияние на морфогенез у борцов, в первую очередь на рост, и в меньшей степени на конституцию. Производные глутаминовой кислоты почти в равной степени связаны с абсолютными и относительными размерами тела.
Что же касается глутаминовой кислоты, по количеству которой борцы являются рекордсменами среди представителей рассмотренных видов спорта, то ее абсолютное количество связано только с относительным.
Корреляционные связи между относительным содержанием аминокислот и морфометрическими параметрами у борцов представлены в таблице 3.2. И здесь преобладают связи с древними аминокислотами, хотя и в меньшей степени (11 связей). И здесь среди древних аминокислот преобладает аспарагиновая кислота.
Разветвленные аминокислоты у борцов не имеют корреляционных связей ни в какой из таблиц.
Какие же морфометрические показатели имеют наибольшее количество корреляций с аминокислотным составом плазмы крови у борцов? Это ширина таза и длина ног, абсолютные показатели. Ширина таза коррелирует с абсолютным количеством аспарагиновой кислоты и пролина, и с относительным содержанием глицина, цистеина и гистидина (все три последние связи — отрицательны). Длина ноги так же имеет 5 корреляционных связей – две с абсолютным количеством аспарагиновой кислоты и пролином, и три с относительным количеством тех же аспарагиновой кислоты и пролина, а также отрицательная связь с относительным количеством цистеина.
Компонентный состав тела (мышечный, костный и жировой компоненты) у борцов имеют корреляционные связи исключительно с древними аминокислотами. Жировой компонент связан с аспарагиновой кислотой и аланином, костный компонент – с аспарагиновой кислотой и глицином. И только мышечный компонент, его относительное к весу тела значение связано с глутаминовой кислотой, и с ее абсолютным и относительным содержанием.
Таблица 3.2 корреляционные связи относительного содержания аминокислот и морфологических показателей у борцов:
| Показатели | «Древние» аминокислоты | Серосодержащие аимнокислоты | Произв. ГЛУ | Разветвленные аминокислоты | Циклические аминокислоты |
| Возраст | АСП, -ГЛИ | ||||
| Рост | -ЦИС | ||||
| ВР | АСП | ||||
| ИП | АСП | ||||
| ШПО | АРГ | ||||
| ШТ | -ГЛИ, | -ЦИС | -ГИС | ||
| ШТО | -ГИС | ||||
| ДРО | -ТРИ | ||||
| ДН | АСП | -ЦИС | ПРО | ||
| ЖК | АСП | ||||
| ЖКО | АСП | ||||
| МКО | ГЛУ | ||||
| ККО | -АСП, ГЛИ | ||||
| ИТОГО 19 | 11 | 3 | 2 | 3 |
Таким образом, отмечаем значение древних аминокислот, а из них – аспарагиновой кислоты для морфометрических показателей борцов. Так же особое положение глутаминовой кислоты в ее связи с мощью мышечного вооружения борца.
Следует также отметить, что повышенное содержание у борцов глутаминовой кислоты и глутамина (Глава 2.Табл.№ 2,3,4,5) не нашло предполагаемого отражения в связях с морфометрическими показателями, за исключением связи содержания глутаминовой кислоты с мышечным компонентом. Пониженное же содержание таурина, орнитина и триптофана (Глава 2.Табл. № 6,7,8,9) нашло отражение в связях с пропорциями тела, шириной плеч и длиной рук.
3.1.2 Корреляции морфометрических показателей с содержанием аминокислот у легкоатлетов
Таблица 3.3 корреляционные связи абсолютного содержания аминокислот и морфологических показателей у легкоатлетов:
| Показатели | «Древние» амнокислоты | Серосоде ржащие | Произв. ГЛУ | Разветвленные
аминокислот |
Циклические
аминокислоты |
| Возраст | -ТИР | ||||
| Рост | СЕР, ГЛИ, АЛА | ||||
| ИП | -АСП, -СЕР, -ГЛИ | -ГЛН | |||
| ДТ | СЕР, ГЛИ, АЛА | ||||
| ДТО | -ВАЛ, -ЛЕЙ | ||||
| ШТ | ВАЛ, ЛЕЙ | ТИР | |||
| ШПО | -ТИР | ||||
| ДР | ГЛИ, АЛА | ГИС | |||
| ДРО | АРГ | ГИС | |||
| ДН | АСП,СЕР, ГЛИ,АЛА | ||||
| ДНО | АСП, СЕР | ИЛЕЙ | |||
| ККО | АСП, АЛА | МЕТ | |||
| ИТОГО 32 | 19 | 1 | 2 | 5 | 5 |
Корреляционные связи абсолютного содержания аминокислот в плазме крови у легкоатлетов с морфометрическими показателями представлены на таблице 3.3. Бросается в глаза обилие связей древних аминокислот – 19. Несколько меньше связей разветвленных и циклических аминокисло – по 5. Здесь уже нет такого засилья со стороны аспарагиновой кислоты – только четыре связи приходится на нее. Обращает на себя внимание то, что абсолютное содержание аспарагиновой кислоты и серина имеют отрицательные связи с индексом пропорциональности и длиной ног, абсолютной и относительной. Иначе говоря, аспарагиновая кислота и серин способствуют проявлению формы тела, специфичной для легкоатлетов.
Корреляционные связи относительного содержания аминокислот в плазме крови легкоатлетов с морфологическими показателями (Табл. 3.4). Преобладание связей древних аминокислот уже не столь заметно – только 7 связей с морфологическими параметрами. Зато резко увеличено количество связей производных глутаминовой кислоты, а именно глутамина.
Таблица 3.4 корреляционные связи относительного содержания аминокислот и морфологических показателей у легкоатлетов:
| Показатели | «Древние» аминокислоты | Серосоде ржащие | Произв. ГЛУ | Разветвленные
аминокислоты |
Циклические
аминокислоты |
| Возраст | ГЛУ, АЛА | -ТРИ | |||
| Рост | -ГЛН | ||||
| Вес | ГЛИ | ||||
| ИП | ГЛН | ||||
| ДТО | ГЛН | ||||
| ШПО | -АСП | ||||
| ШТ | -ГЛН | ||||
| ШТО | ТИР | ||||
| ДРО | -МЕТ | ||||
| ДН | -ГЛН | ||||
| ДНО | АСП | -ФЕН | |||
| ЖК | ЛЕЙ | ||||
| ЖКО | -ГЛИ | ТАУ | ТИР | ||
| ККО | АСП | -ГИС | |||
| ИТОГО 20 | 7 | 2 | 5 | 1 | 5 |
Таким образом, общее количество корреляций между морфологическими параметрами и относительным содержанием аминокислот в плазме крови у легкоатлетов (16) больше, чем у борцов (13). Обращает на себя резкое возрастание корреляций с глутамином, и, особенно, с циклическими аминокислотами. Связей содержания аминокислот с мышечным компонентом у легкоатлетов нет.
Аспарагиновая кислота, серин, глутамин и пролин имеют у легкоатлетов повышенное содержание (Глава 2, табл. №2,3,4,5). Это повышенное содержание для аспарагиновой кислоты, серина и глутамина нашло свое отражение в большом количестве связей с морфологическими параметрами. Содержание же глутаминовой кислоты, таурина метионина и валина у легкоатлетов понижено по сравнению с контролем (Глава 2, табл. №2,3,6,7,8,9). Тем не менее, таурин, метионин и валин образуют единичные в нашем исследовании связи с морфометрическими показателями.
3.1.3 Корреляции морфометрических показателей с содержанием аминокислот у хоккеистов
Таблица 3.5 корреляционные связи АБСОЛЮТНОГО содержания аминокислот и морфологических показателей у ХОККЕИСТОВ:
| Показатели | «Древние» аминокислоты | Серосоде ржащие | Произв. ГЛУ | Разветвленные
аминокислоты |
Циклические
аминокислоты |
| ДТО | ОРН | ||||
| ДРО | -ТАУ | ||||
| ШТ | -ПРО | ||||
| ШТО | -ПРО | ||||
| МК | АСП | ||||
| МКО | -ГЛУ | МЕТ | |||
| ИТОГО 7 | 2 | 2 | 3 |
Представленные на таблице 3.5 корреляционные связи между абсолютным содержанием аминокислот и морфологическими показателями у хоккеистов необычайно скудны – всего семь связей. Две из них приходятся на древние аминокислоты, две приходятся на серосодержащие аминокислоты и три на производные глутаминовой кислоты. Морфологические показатели, связанные с абсолютным содержанием аминокислот у хоккеистов это относительная длина туловища, относительная длина рук, ширина таза абсолютная и относительная и содержание мышечного компонента, так же относительное и абсолютное. Поражает воображение отрицательное значение корреляции между относительным содержанием мышечного компонента и глутаминовой кислотой. У борцов аналогичная корреляционная связь имеет положительное значение.
Совершенно иную картину мы наблюдаем на таблице 3.6. У хоккеистов относительное содержание аминокислот имеет 26 корреляционных связей с морфометрическими параметрами. Так же, как и у борцов и легкоатлетов у хоккеистов наибольшее количество таких связей имеют древние аминокислот (8 связей). Далее в порядке убывания располагаются циклические (6 связей), серосодержащие и производные глутаминовой кислоты (по 5 связей), разветвленные – 2 связи.
У хоккеистов выше контроля содержание цистеина (Глав 2. Табл.№6,7), цистеин с морфометрическими показателями связей не имеет. Ниже контроля у хоккеистов содержание аспарагиновой кислоты, пролина, валина и триптофана. Содержание валина и триптофана не коррелирует с морфометрическими параметрами. Аспарагиновая кислота коррелирует с мышечным компонентом. Содержание пролина коррелирует с шириной плеч и таза, так же, как и содержание глутаминовой кислоты. Интересны взаимоотношения глутаминовой кислоты у хоккеистов с мышечным компонентом. Абсолютное количество глутаминовой кислоты имеет с относительным мышечным компонентом отрицательную связь, а относительное содержание глутаминовой кислоты имеет с этим показателем положительную связь. То есть при сильной выраженности реконструктивного протеолиза (вторая фаза стресса) большое количество глутаминовой кислоты связано с меньшим мышечным компонентом, а при низком уровне реконструктивного протеолиза большое количество глутаминовой кислоты споособствует высокой мышечности хоккеистов.
Таблица 3.6 корреляционные связи относительного содержания аминокислот и морфологических показателей у хоккеистов:
| Показатели | «Древние» аминокислоты | Серо
соде ржащие |
Произв. ГЛУ | Разветвленные
аминокислоты |
Циклические
аминокислоты |
| Возраст | ГЛИ | ЛЕЙ | |||
| Вес | МЕТ | ТИР | |||
| ВР | МЕТ | ТИР | |||
| ИП | ГИС | ||||
| ДТ | ОРН | ||||
| ДТО | ОРН | ГИС | |||
| ШП | ГЛУ | ||||
| ШПО | ПРО | ||||
| ШТ | ГЛУ, | ПРО | |||
| ШТО | ГЛУ, | ПРО | |||
| ДРО | ТАУ | ||||
| ЖК | ГИС | ||||
| МК | АСП, | МЕТ | |||
| МКО | АСП, ГЛУ | ||||
| КК | МЕТ | ТИР | |||
| ККО | ГЛИ | ЛЕЙ | |||
| ИТОГО 26 | 8 | 5 | 5 | 2 | 6 |
Таким образом, выявленная разница в содержании аминокислот в плазме крови спортсменов сопровождается различием в наличии корреляционных связей с морфометрическими показателями. Прежде всего, надо отметить, что наибольшее количество таких связей имеют древние аминокислоты – у представителей всех видов спорта. У борцов и легкоатлетов больше связей абсолютного количества аминокислот, а у хоккеистов наоборот, больше связей с относительным количеством аминокислот.
У хоккеистов выраженность преобладания связей древних аминокислот выражена слабее, чем у борцов и легкоатлетов.
Достаточно отчетливо разница в связях содержания аминокислот и морфометрических параметров видна по отдельным аминокислотам. Так аспарагиновая кислота у борцов преобладает в количестве связей с размерами и пропорциями, у хоккеистов она коррелирует с компонентным составом тела. У легкоатлетов появляются корреляции размеров и пропорций с глутамином, чего нет у представителей других видов спорта. Глутаминовая кислота у борцов связана только с мышечным компонентом, причем положительно, у легкоатлетов – только с возрастом, и у хоккеистов – с пропорциями тела и тоже мышечным компонентом, но одна из связей уже отрицательна.
При выполнении физической работы, в организме происходят изменения функций внутренних органов. Эти изменения у представителей разных видов спорта происходят в соответствии с характером нагрузки и видом деятельности. Подробному исследованию функции сердечно-сосудистой и дыхательной системы посвящено большое количество работ. Но данных о связи особенностей метаболизма аминокислот с особенностями функционирования сердечно-сосудистой и дыхательной систем нет.
3.2 Корреляционные связи между аминокислотным составом плазмы крови и показателями гемодинамики по данным импедансного исследования.
Для выявления механизмов воздействия аминокислот-адаптогенов на физиологические параметры спортсменов был проведен корреляционный анализ динамики количества свободных аминокислот и ряда параметров кардиореспираторнрой системы спортсмена. Подробно результаты исследований кардиореспираторной системы биоимпедансным методом и респираторной системы с помощью традиционных спирометрических методов приведены в ПРИЛОЖЕНИИ. Исследованию состояния кардиореспираторной системы( равно как и морфометрических показателей) у спортсменов различных спецификаций и возрастных групп посвящено огромное количество работ, равно как и механизмам адаптивных перестроек этих систем [14, 15, 17, 18, 48, 60, 62, 63, 64, 80, 107, 108, 109, 112, 114, 154, 155, 156, 169, 171, 173, 174, 193, 194, 195, 196, 197, 198, 260, 242, 261, 283, 288, 306, 307, 308, 309, 310, 311, 313, 314, 319, 320, 321, 322, 323, 324, 325, 326, 327, 328, 339, 338, 376, 378, 352, 353, 354, 355, 356, 366, 386, 387, 388, 389, 390, 391, 395, 53, 57, 495]. Поэтому мы не ставили своей задачей провести подробный сравнительный анализ полученных результатов у исследуемых спортсменов. Здесь же приводятся результаты, которые на наш взгляд позволяют составить некоторое представление о механизмах влияния исследуемых аминокислот на физиологические параметры организма спортсмена.
3.2.1 Корреляционные связи между аминокислотным составом плазмы крови и показателями гемодинамики у борцов
Таблица 3.7 корреляционные связи абсолютного содержания аминокислот и показателей гемодинамики у борцов:
| Показатели | «Древние» аминокислоты | серосодержащие | Произв. ГЛУ | Разветвленные аминокислоты | Циклические аминокислоты |
| PI_ | СЕР, ГЛУ | ||||
| HR_ | |||||
| SpO2 | -ТРИ | ||||
| ToeA | |||||
| NISP | АСП, ГЛИ,-АЛА | -ЦИС | -АРГ | -ЛЕЙ | -ТИР,-ФЕН, -ГИС |
| NIDP | |||||
| @__S_ | |||||
| TrxA | |||||
| SV_ | АРГ | ||||
| EF_ | |||||
| Fw_ | -ГЛУ | ||||
| CO_ | ТИР | ||||
| CI_ | ТИР | ||||
| DO2 | ЦИС | ТИР | |||
| Всего 18 | 6 | 2 | 2 | 1 | 7 |
Наибольшее количество корреляций абсолютного количества аминокислот наблюдается в группе циклических аминокислот и древних. Среди относительного количества аминокислот — в группе производных глутаминовой кислоты и серосодержащих аминокислот.
Интергральный показатель здоровья у борцов имеет положительные корреляции с абсолютным содержанием глутаминовой кислоты и серина (Табл.3.7). Этот показатель у борцов так же связан отрицательными связями с относительным содержанием орнитина, лейцина и тирозина (Табл.3.8).
Частота сердечных сокращений у борцов связана исключительно с относительным содержанием метионина отрицательной корреляционной связью.
Напряжение кислорода в крови достаточно важный показатель. С метаболизмом аминокислот имеет две связи – отрицательную с абсолютным количеством триптофана и положительную с относительным содержанием метионина.
Амплитуда пульсации периферических сосудов и тонус крупных сосудов не связаны с метаболизмом аминокислот у борцов.
Таблица №8 корреляционные связи относительного содержания аминокислот и показателей гемодинамики у борцов:
| Показатели | «Древние» аминокислоты | серосодержащие | Произв. ГЛУ | Разветвленные аминокислоты | Циклические аминокислоты |
| PI_ | -ОРН | -ЛЕЙ | -ТИР | ||
| HR_ | -МЕТ | ||||
| SpO2 | МЕТ | ||||
| ToeA | |||||
| NISP | АСП | ||||
| NIDP | -ГЛИ | -ЦИС | |||
| @__S_ | МЕТ | ||||
| TrxA | |||||
| SV_ | |||||
| EF_ | |||||
| Fw_ | -ГЛУ | ОРН | ВАЛ | ||
| CO_ | -ОРН | -ВАЛ | |||
| CI_ | -ОРН | ||||
| DO2 | -ОРН | ||||
| Всего 16 | 3 | 4 | 5 | 3 | 1 |
Очень многочисленны связи содержания аминокислот и показателей артериального давления. Так систолическое давление связано положительной связью с абсолютным и относительным содержанием аспарагиновой кислоты и отрицательными связями с абсолютным количеством глицина, цистеина, аргинина, лейцина, тирозина, фенилаланина и гистидина. Диастоличесское давление не имеет связей с абсолютным количеством аминокислот. Диастоличесткое давление связано с относительным содержанием глицина и цистеина. Обращают на себя внимание отрицательные связи давления с глицином. Получается, что повышение абсолютного содержания глицина понижает систолическое давление, а повышение относительного содержания глицина понижает диастолическое давление.
Относительное содержание метионина положительно связано с тонусом симпатической нервной системы.
Абсолютное содержание аргинина положительно связано с ударным объемом и отрицательно с систолическим артериальным давлением.
Фракция выброса (эффективность сердечного сокращения) связана отрицательно с относительным содержанием цистеина.
ЦВД отрицательно связано с абсолютным количеством глутаминовой кислоты.
Минутный объем крови и индекс доставки кислорода связаны с абсолютным содержанием тирозина – положительно, и с относительным содержанием орнитина – отрицательно. Таким образом, наиболее сильно влияние аминокислотного состава крови на систолическое давление. Активный реконструктивный протеолиз способствует снижению систолического давления. Исключение составляет аспарагиновая кислота, чья связь с систолическим давлением положительна.
Важнейшая аминокислота у борцов – глутаминовая. Ее количество коррелирует положительно с интегральным показателем здоровья и отрицательно – с центральным венозным давлением. Содержание глутамин у борцов не коррелирует ни с каким показателем гемодинамики. Содержание аминокислот орнитина, таурина и триптофана у борцов ниже, чем в контроле. Содержание таурина не коррелирует с показателями гемодинамики. Содержание орнитина отрицательно связано с показателями тканевого дыхания и интегральным индексом здоровья и положительно – с центральным венозным давлением. И, наконец, содержание триптофана связано с сатурацией кислорода в крови отрицательно.
Абсолютное количество гли положительно связано с систолическим давлением, а относительное – отрицательно связано с диастолическим. То есть глицин повышает емкость сосудов и способность ссс оберспечивать аэробные нагрузки. Поскольку артериальное давление – интегральный показатель, то уровень глицина повышая размах А/Д у борцов является показателем способности эфффективно обеспечивать усиление кровоообращения при физической нагрузке. Никакой адреналин по степени регуляторных возможностей у борцов в отношении глицина не сопоставим по регуляторным возможностям.
Кроме того, на показатели артериального давления влияет и абсолютно и относительно цистеин, в обратно пропорциональной зависимости.
На показатели ВНС положительно влияет метионин. Он положительно повышает тонус симпатики у борцов. Чем меньше цистеина, тем больше фракция выброса. Уровень цистеина неметаболизируемый в крови отрицательно влияет на способность организма эффективно обеспечивать организм при тонических нагрузках. Лишний цистеин борцам категорически не нужен, поэтому с ростом мастерства у борцов абсолютное количество цистеина снижается в полтора раза (р<0,01), и есть тенденция к снижению относительного количества цистеина. Прямая связь абсолютного содеожания цистеина с индексом доставки кислорода.
Высокие показатели цистеина говорят о излишне высоком уровне метаболизма в организме борца.
3.2.2 Корреляционные связи между аминокислотным составом плазмы крови и показателями гемодинамики у легкоатлетов
Таблица 3.9 корреляции между показателями гемодинамики и АБСОЛЮТНЫМ содержанием аминокислот в плазме крови у ЛЕГКОАТЛЕТОВ:
| Показатели | «Древние» аминокислоты | серосодержащие | Произв. ГЛУ | Разветвленные аминокислоты | Циклические аминокислоты |
| PI_ | -ЦИС | -ТРИ | |||
| HR_ | МЕТ | ||||
| SpO2 | -ЦИС | ||||
| ToeA | АСП, СЕР | ||||
| NISP | -ТАУ | -ФЕН | |||
| NIDP | |||||
| @__S_ | -ТАУ | -ФЕН | |||
| TrxA | |||||
| SV_ | |||||
| EF_ | -МЕТ | ||||
| Fw_ | ИЛЕ | ||||
| CO_ | |||||
| CI_ | |||||
| DO2 | |||||
| Всего 12 | 2 | 6 | 1 | 3 |
Изучение корреляций между показателями гемодинамики и абсолютным содержанием аминокислот в плазме крови у легкоатлетов (Табл. 3.9) позволяет установить, что наибольшее количество связей с показателями гемодинамики дают серосодержащие аминокислоты (6). Древние аминокислоты – аспарагиновая и серин положительно связаны с амплитудой пульсации капилляров.
Представленные нами ниже корреляции между показателями гемодинамики и относительным (Табл. 3.10) содержанием аминокислот в плазме крови у легкоатлетов позволяют увидеть, что наибольшее количество корреляций приходится на циклические аминокислоты. Иначе говоря, серосодержащие аминокислоты оказывают влияние метаболическое, а циклические аминокислоты – регуляторное.
Таблица 3.10 корреляции между показателями гемодинамики и ОТНОСИТЕЛЬНЫМ содержанием аминокислот в плазме крови у ЛЕГКОАТЛЕТОВ:
| Показатели | «Древние» аминокислоты | серосодержащие | Произв. ГЛУ | Разветвленные аминокислоты | Циклические аминокислоты |
| PI_ | -ТРИ | ||||
| HR_ | |||||
| SpO2 | -ТРИ | ||||
| ToeA | -ФЕН | ||||
| NISP | -ТАУ | ||||
| NIDP | |||||
| @__S_ | -ТАУ | ||||
| TrxA | |||||
| SV_ | -ТРИ | ||||
| EF_ | -АСП,-СЕР | -ГИС | |||
| Fw_ | -ГЛУ | ||||
| CO_ | |||||
| CI_ | |||||
| DO2 | -ТРИ | ||||
| Всего 11 | 3 | 3 | 5 |
Связи интегрального показатели здоровья говорят о том, что этот показатель повышается при повышении абсолютного содержания метионина. Повышение абсолютного содержания цистеина и абсолютного и относительного содержания триптофана понижают интегральный показатель здоровья.
ЧСС у легкоатлетов положительно связана с абсолютным содержанием метионина.
Содержание кислорода ограничено содержанием цистеина (абсолютным) и триптофана (относительным) – между ними связи отрицательны.
АПФГ связана исключительно с абсолютным содержанием всех древних аминокислот, причем с глутаминовой связь – отрицательна.
Тонус симпатической нервной системы отрицательно связан с абсолютным и относительным содержанием таурина, и отрицательно же с относительным содрержанием фенилаланина.
Ударный объем имеет единственную отрицательную связь с относительным содержанием триптофана.
Фракция выброса, как показатель эффективности работы сердца имеет множество связей. Отрицательная связь с абсолютным содержанием метионина дополнена отрицательной связью с относительным содержанием аспарагиновой кислоты, серина и положительной – с гистидином.
Таким образом, преобладающие количественно аспарагиновая кислота и серин имеют положительные связи с амплитудой пульсации капилляров и отрицательные связи с эффективностью работы сердца. Эти две аминокислоты, таким образом, способствуют микроциркуляции при высоком уровне реконструктивного протеолиза.
Из двух других аминокислот, чье содержание у легкоатлетов повышено – пролин и глутамин, ни одна не имеет корреляционных связей с показателями гемодинамики, при том, что глутамин имеет много связей с размерами и пропорциями тела у легкоатлетов.
Содержание серосодержащих аминокислот таурина и метионина у легкоатлетов понижено. Содержание таурина отрицательно связано с систолическим давлением и отрицательно — с тонусом симпатической нервной системы. Метионин – повышает частоту сердечных сокращений и снижает фракцию выброса.
3.2.3 Корреляционные связи между аминокислотным составом плазмы крови и показателями гемодинамики у хоккеистов
Таблица 3.11 корреляции между показателями гемодинамики и АБСОЛЮТНЫМ содержанием аминокислот в плазме крови у ХОККЕИСТОВ:
| Показатели | «Древние» аминокислоты | серосодержащие | Произв. глу | Разветвленные аминокислоты | Циклические аминокислоты |
| PI_ | |||||
| HR_ | |||||
| SpO2 | СЕР, ГЛИ | ГЛН | |||
| ToeA | АРГ | ||||
| NISP | |||||
| NIDP | -АСП | ||||
| @__S_ | -СЕР, -ГЛИ | -ГЛН | -ЛЕЙ | -ФЕН | |
| TrxA | ГЛИ | ЦИС | |||
| SV_ | СЕР,-ГЛУ | ||||
| EF_ | |||||
| Fw_ | |||||
| CO_ | -ГЛУ | ||||
| CI_ | -ГЛУ | ||||
| DO2 | -ГЛУ | ||||
| Всего 17 | 11 | 1 | 3 | 1 | 1 |
Интересно распределение корреляционных связей между содержанием аминокислот и показателями гемодинамики у хоккеистов. Абсолютное содержание древних аминокислот имеет большое количество связей с показателями гемодинамики, а относительное содержание древних аминокислот у хоккеистов не имеет ни одной такой связи (Табл.3.11 и 3.12).
Интегральный показатель здоровья у хоккеистов связан только с относительным содержанием метионина. Частота сердечных сокращений имеет отрицательную связь с триптофаном.
Сатурация кислорода в крови имеет положительную связь с абсолютным содержанием серина, глицина и глутамина и относительным содержанием глутамина.
Амплитуда пульсации капилляров положительно связана с содержанием аргинина, абсолютным и относительным.
Артериальное давление имеет единственную корреляционную связь у хоккеистов – отрицательную связь абсолютного содержания аспарагиновой кислоты с диастолическим давлением.
Высокое абсолютное содержание пяти аминокислот способствуют снижению тонуса симпатической нервной системы. Это серин, глинцин, глутамин, лейцин и фенилаланин. Таким образом высокое абсолютное содержание этих аминокислот снижает эффект первой фазы стресса. Относительное содержание этих аминокислот таких связей не имеет.
Тонус крупных сосудов положительно связан с абсолютным содержанием глицина и цистеина и отрицательно с относительным содержанием цистеина.
Ударный объем положительно связан с абсолютным содержанием серина, и имеет две отрицательные связи – с абсолютным содержанием глутаминовой кислоты и относительным содержанием валина.
Эффективность сердечных сокращений и центральное венозное давление у хоккеистов с метаболизмом аминокислот связей не имеет.
Эффективность кровообращения отрицательно связано с абсолютным содержанием глутаминовой кислоты.
Цистеин – единственная аминокислота у хоккеистов, содержание которой выше, чем в контроле (Глава 2, табл. №2). Связь только с тонусом крупных сосудов.
Четыре аминокислоты у хоккеистов имеют содержание ниже, чем в контроле. Это аспарагиновая кислота, пролин, валин и триптофан. Содержание пролина с показателями гемодинамики связей не имеет, аспарагиновая кислота связана с диастолическим давлением, валин – с тонусом крупных сосудов и триптофан – с частотой сердечных сокращений.
Таблица 3.12 корреляции между показателями гемодинамики и ОТНОСИТЕЛЬНЫМ содержанием аминокислот в плазме крови у ХОККЕИСТОВ:
| Показатели | «Древние» аминокислоты | Серосодержащие | Произв. ГЛУ | Разветвленные аминокислоты | Циклические аминокислоты |
| PI_ | МЕТ | ||||
| HR_ | -ТРИ | ||||
| SpO2 | ГЛН | ||||
| ToeA | АРГ | ГИС | |||
| NISP | |||||
| NIDP | |||||
| @__S_ | |||||
| TrxA | -ВАЛ | ||||
| SV_ | -АРГ | ||||
| EF_ | |||||
| Fw_ | |||||
| CO_ | |||||
| CI_ | |||||
| DO2 | |||||
| Всего 7 | 1 | 3 | 1 | 2 |
3.3 Корреляционные связи показателей внешнего дыхания и аминокислотного состава плазмы крови у спортсменов
3.3.1 Корреляционные связи между аминокислотным составом плазмы крови и показателями внешнего дыхания у борцов
Дыхательные объемы измерялись с помощью спирометра, определялись резервные объемы вдоха и выдоха, жизненная емкость легких фактическая, по данным морфометрии – расчетная жизненная емкость легких, максимальный дыхательный объем при нагрузочном тестировании, минутный объем дыхания максимальный, и удельный дыхательный объем (на ватт энергии), а так же ПВЦ170, удельный ПВЦ170, и критическая удельная мощность (удельная мощность при которой снижается эффективность дыхательной функции).
Резервный объем вдоха и резервный объем выдоха у борцов имеет две группы связей. Ряд связей имеет реципрокный характер. Так абсолютное количество серина и глутамина имеет положительную связь с резервным объемом вдоха и отрицательную – с резервным объемом выдоха. Кроме того, относительное количество изолейцина имеет отрицательную связь с резервным объемом вдоха и положительную – с резервным объемом выдоха. К числу других связей относятся отрицательные связи резервного объема выдоха с абсолютным содержанием глицина и триптофана и положительная связь с лейцином (Табл. 3.13, 3.14).
Фактическая ЖЭЛ связана отрицательной связью с абсолютным содержанием орнитина и триптофана и относительным содержанием триптофана же.
Расчетная ЖЭЛ не имеет связей с метаболизмом аминокислот.
Таблица 3.13 корреляционные связи абсолютного содержания аминокислот и показателей дыхательной функции у борцов:
| Показатели | «Древние» аминокислоты | Серосодержащие | Произв. ГЛУ | Разветвленные аминокислоты | Циклические аминокислоты | Всего |
| Ровд | СЕР | ГЛН | 2 | |||
| Ровыд | -СЕР,-ГЛИ | -ГЛН | -ТРИ | 4 | ||
| Жэлфакт | -ОРН | -ТРИ | 2 | |||
| Жэлрасч | ||||||
| ДОмакс | -ЦИС | ГЛН | 2 | |||
| МОДмакс | ГЛН | 1 | ||||
| удДОмакс | ||||||
| пвц170 | ||||||
| Пвцудель | АСП | 1 | ||||
| Кум | ||||||
| 4 | 1 | 5 | 2 | 12 |
Максимальный дыхательный объем и максимальный минутный объем у борцов связаны с абсолютным содержанием глутамина. Абсолютное содержание глутамина в крови у борцов, таким образом, связано с эффективностью функции внешнего дыхания. Возможность дышать у борцов поддержана интенсивностью реакций детоксикации в головном мозге. Содержание цистеина имеет отрицательную связь с максимальным дыхательным объемом, то есть имеет больше связи со структурой органов дыхания, чем с функцией. Относительное содержание лейцина тоже связано с максимальным дыхательным объемом и с минутным объемом, по- разному. С максимальным дыхательным объемом эта связь положительна, а с минутным дыхательным объемом – отрицательна, так же как и с удельным дыхательным объемом.
Мощность при тестировании PWC170 у борцов не связана с аминокислотным составом плазмы крови. При этом удельная мощность положительно связана с содержанием аспарагиновой кислоты и отрицательно – с относительным содержанием аланина.
И, наконец, критическая удельная мощность (при которой происходит срыв дыхания) связана отрицательно с относительным содержанием лейцина.
Таким образом, можно сказать, что разветвленные аминокислоты (относительное их содержание) у борцов оказывают всестороннее влияние на функцию внешнего дыхания.
Таблица 3.14 корреляционные связи относительного содержания аминокислот и показателей дыхательной функции у борцов:
| Показатели | «Древние» аминокислоты | Серосодержащие | Произв. ГЛУ | Разветвленные аминокислоты | Циклические аминокислоты | Всего |
| Ровд | -ИЛЕ | 1 | ||||
| Ровыд | -ГЛН | ИЛЕ, ЛЕЙ | 3 | |||
| Жэлфакт | -ТРИ | 1 | ||||
| Жэлрасч | ||||||
| ДОмакс | ЛЕЙ | 1 | ||||
| МОДмакс | -ЛЕЙ | 1 | ||||
| удДОмакс | -ЛЕЙ | 1 | ||||
| пвц170 | ||||||
| Пвцудель | АСП, -АЛА | 2 | ||||
| Кум | -ЛЕЙ | 1 | ||||
| 2 | 1 | 6 | 1 | 10 |
3.3.2 Корреляционные связи между аминокислотным составом плазмы крови и показателями внешнего дыхания у хоккеистов.
Таблица 3.15 корреляционные связи абсолютного содержания аминокислот и показателей дыхательной функции у ХОККЕИСТОВ:
| Показатели | «Древние» аминокислоты | Серосодержащие | Произв. ГЛУ | Разветвленные аминокислоты | Циклические амитнокислоты | Всего |
| Ровд | -МЕТ | -ИЛЕ | 2 | |||
| Ровыд | МЕТ | ИЛЕ | 2 | |||
| Жэлфакт | АСП | 1 | ||||
| Жэлрасч | -ОРН,-АРГ | -ГИС | 3 | |||
| ДОмакс | ||||||
| МОДмакс | -ГЛУ, ГЛИ | 2 | ||||
| удДОмакс | -ГЛУ | 1 | ||||
| пвц170 | ||||||
| Пвцудель | ТИР | 1 | ||||
| Кум | ГЛИ | -ТАУ | 2 | |||
| 5 | 3 | 2 | 2 | 2 | 14 |
Резервные объемы вдоха и выдоха у хоккеистов, так же, как и у борцов, имеют корреляционные связи с абсолютным содержанием аминокислот реципрокного характера (Табл.3.15, 3.16). В отличие от борцов отрицательные связи абсолютного содержания метионина и изолейцина у хоккеистов с резервным объемом вдоха, а положительные – с резервным объемом выдоха.
Фактическая жизненная емкость легких у хоккеистов имеет положительную корреляционную связь с абсолютным содержанием аспарагиновой кислоты. Расчетная жизненная емкость легких имеет отрицательные корреляционные связи с абсолютным содержанием орнитина, аргинина и гистидина и положительную связь с относительным содержанием тирозина.
Максимальный дыхательный объем ухоккеистов не имеет корреляционных связей с показателями метаболизма аминокислот. Минутный же максимальный объем дыхания у хоккеистов положительно связан с абсолютным содержанием глицина и отрицательно – с абсолютным содержанием глутаминовой кислоты. Этот же показатель связан отрицательно с относительным содержанием серина и метионина и положительно – с относительным содержанием глутамина.
Удельный дыхательный объем (на единицу мощности работы) у хоккеистов связан с метаболизмом аминокислот. Это отрицательная связь с абсолютным содрежанием глутаминовой кислоты, отрицательная связть с относительным содержанием метионина и положительная с относительным содержанием орнитина.
Мощность на пульсе 170 у хоккеистов с метаболизмом аминокислот не имеет связей. Удельная мощность на пульсе 170 (на килограмм веса) связана с абсолютным содержанием тирозина.
Критическая удельная мощность у хоккеистов связана положительно с абсолютным содержанием глицина и отрицательно с абсолютным содержанием таурина. Кроме того – с относительным содержанием глутамина и аргинина, положительно.
Таблица 3.16 корреляционные связи относительного содержания аминокислот и показателей дыхательной функции у ХОККЕИСТОВ:
| Показатели | «Древние» аминокислоты | Серо-содержащие | Произв. ГЛУ | Разветвленные аминокислоты | Циклические аминокислорты | Всего |
| Ровд | ||||||
| Ровыд | ||||||
| Жэлфакт | -ГЛИ | 1 | ||||
| Жэлрасч | ТИР | 1 | ||||
| ДОмакс | ||||||
| МОДмакс | -СЕР | -МЕТ | ГЛН | 3 | ||
| удДОмакс | -МЕТ | ОРН | 2 | |||
| пвц170 | ||||||
| Пвцудель | ||||||
| Кум | ГЛН, АРГ | 2 | ||||
| 2 | 2 | 4 | 1 | 9 |
3.4 ПСИХОЛОГИЯ Корреляционные связи между аминокислотным составом плазмы крови спортсменов и их психологическими параметрами.
С целью выяснения вопроса о взаимозависимости метаболизма и психологических характеристик было проведено психологическое тестирование спортсменов. Исследовались – темперамент по Айзенку, показатели психологической соревновательной устойчивости по Мильману, самооценка спортсменов с использованием теста САН и для оценки склонности к формированию фиксированных форм поведения, как степень патологичности приспособления к деятельности тест ТОРЗ.
Исследование проводилось по стандартной методике:
Данные по психологическому тестированию приведены в приложении. В данном исследовании приведены показатели корреляционных связей между психологическими показателями и аминокислотным составом. Даны средние показатели для группы.
3.4.1 Корреляционные связи показателей темперамента и содержанием аминокислот в плазме крови спортсменов.
3.4.1.1. Борцы
Данные по исследованию темперамента представлены в ПРИЛОЖЕНИИ. Здесь мы рассматриваем корреляционные связи показателей темперамента и содержанием аминокислот в плазме крови у спортсменов.
У борцов корреляционных связей между темпераментом и древними аминокислотами (абсолютно и относительно) в крови нет (Табл.3.17).
Таблица 3.17 сводные данные о корреляционных связях между показателями темперамента и содержанием аминокислот в плазме крови у борцов:
| Показатели | «Древние» амнокислоты | Серосоод | Произв. ГЛУ | Разветвленные аминокислоты | Циклические аминокислоты |
| Абсолютное содержание аминокислот | |||||
| Экстраверсия | |||||
| Нейротизм | |||||
| Относительное содержание аминокислот | |||||
| Экстраверсия | — ВАЛ | ||||
| Нейротизм | ЦИС | ОРН | |||
Экстраверсия есть функция, связанная с формированием афферентного синтеза пространственного поля, иначе говоря, экстраверсия есть функция активности правого полушария.
Нейротизм есть отрицательный показатель контроля за эмоциями, или, иначе говоря, есть показатель активности левого полушария, точнее, его неактивности (чем ниже нейротизм, тем выше активность левого полушария).
Орнитин у борцов отрицательно влияет на ИДК и на кровообращение, и здесь видим его положительное влияние на нейротизм, то есть подавляет функцию левого полушария по сравнению с правым. О том же говорит положительная связь между нейротизмом и цистеином. Цистеин, как и орнитин отрицательно влияет на ИДК. Цистеин понижает фракцию выброса. Кроме того, высокий уровень цистеина говорит об излишне высоком уровне метаболизма у борцов. С ростом квалификации уровень цистеина у борцов понижается. Таким образом, у борцов аминокислоты, ответственные за детоксикацию снижают функцию левого полушария. Известно, что в головном мозге имеются ферменты цикла биосинтеза мочевины [22,24], в ходе этого процесса может образовываться большое количество орнитина, хотя полностью орнитиновый цикл в мозге до конца не идет.
Что касается серосодержащих аминокислот, цистеина, цистотионина, изотеиновой кислоты, цистеиновой кислоты, как и таурина, они находятся в мозговой ткани в большом количестве, что было ранее непонятно [22], то в свете наших данных их значение становится понятным. Возможно, это связано с регуляцией функции мембран нейронов.
Отрицательная связь между относительным содержанием в крови валина и экстраверсией у борцов – валин понижает функцию правого полушария. Абсолютное количество валина у борцов с ростом квалификации снижается, а относительное количество – повышается. Такая направленность изменений с ростом квалификации может быть связана с усилением анаболических процессов и возможным повышением регуляторной роли валина в метаболизме.
3.4.1.2 Легкоатлеты
Таблица 3.18 сводные данные о корреляционных связях между показателями темперамента и содержанием аминокислот в плазме крови у легкоатлетов:
| Показатели | «Древние» аминокислоты | Серосоодержащие | Произв. ГЛУ | Разветвленные аминокислоты | Циклические аминокислоты |
| Абсолютное содержание аминокислот | |||||
| Экстраверсия | АЛА | ЦИС | |||
| Нейротизм | |||||
| Относительное содержание аминокислот | |||||
| Экстраверсия | -ИЛЕ | ||||
| Нейротизм | |||||
Данные представленные на таблице 3.18 позволяют увидеть, что нейротизм, то есть степень вольности лимбики (недостаточность левого полушария для управления лимбической системой) у легкоатлетов не имеет связей метаболизмом аминокислот. Экстраверсия, то есть степень преобладания активности правого полушария связана положительно с абсолютным количеством аланина и цистеина и отрицательно – с относительным содрежанием изолейцина.
3.4.1.3 Хоккеисты
Относительное содержание в крови у хоккеистов таурина связано отрицательной связью с экстраверсией (Табл.3.19), то есть таурин, будучи стабилизатором мембран снижает функциональное значение правого полушария. Принимая во внимание, что из рассмотренных видов спорта хоккеисты имеют самое высокое абсолютное и относительное содержание таурина, отметим, что функция правого полушария у хоккеистов нуждается в метаболическом контроле, этот контроль осуществляется с помощью таурина. С другой стороны, с ростом квалификации уровень таурина снижается, то есть контроль за поведением переходит на другие, не метаболические уровни.
Таблица 3.19 сводные данные о корреляционных связях между показателями темперамента и содержанием аминокислот в плазме крови у хоккеистов:
| Показатели | «древние» аминокислоты | Серосоодержащие | Произв. ГЛУ | Разветвленные аминокислоты | Циклические аминокислоты |
| Абсолютное содержание аминокислот | |||||
| Экстраверсия | |||||
| Нейротизм | |||||
| Относительное содержание аминокислот | |||||
| Экстраверсия | -ТАУ | ||||
| Нейротизм | -ВАЛ | -ФЕН | |||
Относительное содержание валина и фенилаланина тоже отрицательно связано с нейротизмом, то есть повышают функциональное значение левого полушария. С ростом мастерства содержание этих аминокислот у хоккеистов снижается.
Обращает на себя внимание, что при сравнении с борцами, у которых корреляции между показателями нейротизма и аминокислотами (орнитин и цистеин) связаны положительными связями, то есть повышение их относительной концентрации повышает уровень нейротизма, или, иначе говоря снижает контроль за лимбикой левого полушария. Таким образом аминокислоты – регуляторы нейротизма у борцов и у хоккеистов действуют разнонаправленно. У борцов – снижают, а у хоккеистов – повышают контроль левого полушария за лимбикой. Мы полагаем, что это связано с особенностями деятельности борцов и хоккеистов. Борцы действуют индивидуально, и их эмоциональное состояние касается только лично их и их противников. А хоккеисты действуют в команде, для них важно согласование действий с другими членами команды, и их метаболизм помогает им в этом, при повышении названных метаболитов усиливается сознательный контроль за поведением.
Таурин – фактор общетормозный, фала – предшественник гормонов, если его много, то гормоны не образуются. Валин как неспецифический метаболит, связанный с разнообразием метаболических процессов повышает возможности разнообразия реакций левого полушария.
У хоккеистов корреляционных связей между абсолютным и относительным содержанием древних аминокислот в крови и показателями темперамента нет.
3.4.2. КОРРЕЛЯЦИОННЫЕ СВЯЗИ ПОКАЗАТЕЛЕЙ САМООЦЕНКИ, АКТИВНОСТИ, НАСТРОЕНИЯ (САН) С СОДЕРЖАНИЕМ АМИНОКИСЛОТ В ПЛАЗМЕ КРОВИ СПОРТСМЕНОВ
3.4.2.1 Борцы
Таблица 3.20 сводные данные о корреляционных связях между показателями САН и содержанием аминокислот у борцов:
| Показатели | «Древние» аминокислоты | Серосодержащие | Произв. ГЛУ | Разветвленные аминокислоты | Циклические аминокислоты |
| Абсолютное содержание аминокислот | |||||
| Самочувствие | -ТРИ | ||||
| Активность | |||||
| Настроение | |||||
| Относительное содержание аминокислот | |||||
| Самочувствие | -ТРИ | ||||
| Активность | |||||
| Настроение | ГЛУ, -ГЛИ | -ГЛН | -ТРИ | ||
Самооценка состояния у борцов при тестировании САН имеет 5 связей с относительным содержанием аминокислот и одну связь с абсолютным количеством аминокислот (Табл.3.20). Количество триптофана абсолютное и относительное отрицательно влияет на самочувствие, ничего удивительного в этом нет, поскольку триптофан метаболизируется в серотонин (антистрессовый фактор) а во -вторых – в нуклеотиды дыхательных ферментов. а на настроение отрицательно влияет только относительное количество триптофана. Значительное падение уровня триптофана у борцов с ростом квалификации говорит о том, что для них важнейшее значение имеет расход триптофана с образованием его метаболитов, в данном случае речь идет, скорее всего, о серотонине. Избыток триптофана снижает самочувствие и настроение, но на настроение влияют еще и глутамин, глутаминовая кислота и глицин. Количество триптофана, глутамина и глицина с ростом мастерства у борцов снижаются (р<0,01), и триптофан на самочувствие, а глицин, глутамин и триптофан на настроение своим избытком влияют отрицательно. Глутаминовая кислота, являясь главным метаболитом борцов, влияет на настроение положительно, а с ростом мастерства ее количество у борцов растет, абсолютно и относительно.
Глутамин отрицательно влияет потому, что это метаболит процессов детоксикации аммиака, как в мышцах, так и в мозге. Повышенное его количество в крови говорит о недостаточности печени и почек, то есть о недостаточности детоксикационных процессов.
Глицин – общий тормозный медиатор, и для борца его большие концетрации в крови нежелательны.
Что касается глютаминовой кислоты – этот возбуждающий медиатор ЦНС и антигикопксант и антиоксидант, фактор стимуляции анаболических процессов, стимулирует СТГ (Генкин и Волков и Глотов). Влияет на гормоны щитовидной железы, оптимизируя окислительное-восстановительные процессы. Поэтому характер нагрузки борцов делает глютаминовую кислоту универсальным метаболитом для такого вида спорта.
3.4.2.2 Легкоатлеты
Таблица 3.21 сводные данные о корреляционных связях между показателями САН и содержанием аминокислот у легкоатлетов:
| Показатели | «Древние» аминокислоты | Серосодержащие | Произв. ГЛУ | Разветвленные аминокислоты | Циклические аминокислоты |
| Абсолютное содержание аминокислот | |||||
| Самочувствие | -МЕТ | ||||
| Активность | ГЛН, ПРО | ТРИ | |||
| Настроение | |||||
| Относительное содержание аминокислот | |||||
| Самочувствие | ГИС | ||||
| Активность | -МЕТ | ТРИ | |||
| Настроение | |||||
Глицин – второй маркер легкоатлетов. При вариационном анализе установлено, что уровень глицина в крови у легкоатлетов имеет самое стабильное содержание, хотя в других группах он сильно колебался ( Табл 3.21). Глицин является универсальным тормозным медиатором периферической нервной системы. Что необходимо для выработки двигательных стереотипов. У некоторых насекомых в тканях глицин содержится в огромном количестве [357]. Глицин – донатор метельных групп, что важно для поддержания мембранных структур, так и для образования биологически активных медиаторов (фолиевая кислота, биосинтез нуклеотидов и других БАВ).
Аргинин, как нейропротектор защищает нейроны от чрезмерных стрессовых воздействий, как физических так и психологичческих. В отличие от борцов, у которых начальный (анаэробный) этап работы обеспечен глутаминовой кислотой и мышечным гликогеном, у легкоатлетов есть необходимость такой защиты за счет аргинина.
Отрицательная связь активности с тирозином говорит о зависимости активности легкоатлетов от адреналовых гормонов. Опять же в отличие от борцов, у которых активность обеспечена глутаминовой кислотой, обеспечивающей им лабильность мембран и подвижность нервных процессов.
Глутамин – маркер мозговика и легкоатлета. Много глутамина – значит недостаточно мощные анаболические и детоксикационные процессы в печени, поэтому при снижении количества глутамина в крови – организм очищается от избытка метаболитов, но привычка иметь повышенный глутамин связывает настроение легкоатлета с улучшением настроения при его повышении.
3.4.2.3 Хоккеисты
Таблица 3.22 сводные данные о корреляционных связях между показателями САН и содержанием аминокислот у ХОККЕИСТОВ:
| Показатели | «Древние» аминокислоты | Серосодержащие | Произв. ГЛУ | Разветвленные аминокислоты | Циклические аминокислоты |
| Абсолютное содержание аминокислот | |||||
| Самочувствие | -АРГ | ||||
| Активность | |||||
| Настроение | |||||
| Относительное содержание аминокислот | |||||
| Самочувствие | -ТАУ | ||||
| Активность | -ТАУ | ГИС | |||
| Настроение | ВАЛ | ФЕН, ГИС | |||
Асолютное содержание аргинина и относительное содержание таурина у хоккеистов имеют отрицательные связи с самочувствием. Относительное содеражине таурина связано с активностью отрицательной связью, а относительное содержание гистидина связано с активностью положительно. Относительное содержание валина, фенилаланина и гистидина способствуют хорошему настроению хоккеистов. Понижение активности вполне естественно для таурина, как стабилизатора мембран, но понижение самочувствия ограничивает излишнюю его выработку у хоккеистов (Табл. 3.22).
Значение таурина для экстраверсии (функционального значения правого полушария) у хоккеистов – снижение экстраверсии, но также отрицательно относительное содержание таурина в крови у хоккеистов влияет и на самовчуствие и на активность. Высокое содержание таурина снижает самочувствие, а кроме того и настроение. В отношении активности антагонистом для таурина у хоккеистов является гистидин – связан с активностью по самооценке положительно.
Таурин – тормозный медиатор, и небольшое его снижение для них кайф. Он же снижает активность, а гистидин – медиатор подвижности реакций.
Особенность хоккеистов привыкли в состоянии высокой нагрузки и
Содержание производных глутаминовой кислоты отрицательно связано с настроением. А именно – с абсолютным содержанием аргинина, и относительным содержанием валина и фенилаланина. Эти аминокислоты отрицательно влияют на уровень нейротизма, то есть усиливают левополушарный контроль за лимбической системой, вследствие чего вполне естественно положительно влияют на настроение [296].
Аргинин является протектором по отношению к нейронам, и не позволяет спортсмену слишком раскрепощаться. В силу особенностей тренировочного режима он оказывает отрицательное влияние на самочувствие.
Повышение количества фенилаланина говорит о том, что избыток гормонов им тоже не нужно для настроения. Им нужен для настроения валин – разнообразие тонкой метаболической регуляции метаболизма- выйти из тренировочного состояния – повышает настроение.
Корреляционные связи между САН и абсолютным и относительным содержанием «древних» аминокислот у хоккеистов отсутствуют.
3.4.3. Корреляционные связи оказатели соревновательной устойчивости (по Мильману) с содержанием аминокислот в плазме крови спортсменов
3.4.3.1 Борцы
Всего показатели соревновательной устойчивости у борцов имеют 8 корреляционных связей с относительным количеством отдельных аминокислот, и одну связь с абсолютным количеством глутаминовой кислоты. Обращает на себя внимание, что связи преимущественно отрицательные (6 из 8). Мотивационно-энергетический компонент (МЭ) снижается при относительном повышении орнитина, тирозина и гистидина, но повышен при относительном и, особенно, при абсолютном повышении содержания глутаминовой кислоты.
Эмоциональная устойчивость (СЭУ) понижается при повышенном относительном содержании цистеина и гистидина, но относительно повышенное содержание серина повышает соревновательную эмоциональную устойчивость.
Саморегуляция (СР) у борцов отрицательно связана с относительным повышением глутаминовой кислоты.
Таблица 3.23 сводные данные о корреляционных связях между показателями СОРЕВНОВАТЕЛЬНОЙ УСТОЙЧИВОСТИ (ПО МИЛЬМАНУ) и содержанием аминокислот у БОРЦОВ:
| Абсолютное содержание | «Древние» аминокислоты | Серосодержащие | Произв. ГЛУ | Разветвленные аминокислоты | Циклические аминокислоты |
| Сэу | |||||
| Ср | |||||
| Мэ | ГЛУ | ||||
| Стп | |||||
| Относительное содержание | |||||
| Сэу | СЕР | -ЦИС | -ГИС | ||
| Ср | -ГЛУ | ||||
| Мэ | ГЛУ | -ОРН | -ТИР, -ГИС | ||
| Стп |
На стабильность результатов в соревнованиях (СТП) у борцов метаболизм аминокислот напрямую не влияет.
На эмоциональную устойчивость (СЭУ) у борцов положительно влияет относительное содержание в крови серина, отрицательно – относительное содержание цистеина и гистидина. Про серин можно сказать только то, что его содержание с ростом мастерства снижается абсолютно и относительно (Табл. 3.23).
Серин – как предшественник серосодержащих амнокислот, фосфолипидов мембран мембран положительно влияет на эмоциональную устойчивость.
Цистеин – сам на нее не влияет, а его производные – влияют (таурин и глутатион) поэтому если его много, то мало производных. То есть он не матаболизируется до отмеченных метаболитов.
Гистидин, как и цистеин, должен быстро переходить в свои производные, которые выполняют медиаторную функцию(гистамин), поэтому если его много, то стабильность понижается.
Относительное содержание цистеина и гистидина отрицательно связано с эмоциональной устойчивостью. Избыток цистеина и гистидина не допускают мобильной регуляции энергетического обмена, в результате чего борцам необходимо дополнительное эмоциональное воздействие, что лишает их эмоциональной стабильности.
Глутаминовая кислота, ее абсолютное и относительное содержание связано положительно с мотивацией и мотивационной энергетикой (МЭ). Это вполне естественно, так как постоянные гипоксические нагрузки у борца формируют постоянную же потребность в глутаминовой кислоте, и высокое ее содержание само по себе становится фактором хорошего настроения борца с одной стороны, а с другой стороны вызывает потребность в нагрузках, характерных для борца – сопровождающихся гипоксией, статических усилиях и скоростно-силовых нагрузках. Кроме того, глутаминовая кислота является фактором, снижающим саморегуляцию, положительной стороной чего является высокая переключаемость в экстремальных условиях.
На мотивацию и ее эмоциональную энергетику кроме глутаминовой кислоты влияют орнитин, тирозин и гистидин, причем в отличие от глутаминовой кислоты они влияют отрицательно. Высокий метаболический уровень, недостаточная функция детоксикации и недостаточность катехоламинов – все это мешает быть борцу собранным и быть готовым к бою.
3.4.3.2 Хоккеисты
Влияние аимнокислот на СЭУ. Глицин как тормозный медиатор естественно способствует эмоциональной устойчивости. Валин – интегратор обменных процессов тоже способствует устойчивости. При повышении количества тирозина – меньше образуется адреналовых и тиреоидных гормонов, так же, как и фенилаланина. Увеличение количества валина, лейцина и изолейцинаейцина может свидетельствовать о том, что чем разветвленных аминокислот больше в крови, тем сильнее стрессовая реакция и наоборот.
Сила эмоциональной мотивации корелирует с уровенем изолейцина – то есть с силой стрессовых метаболических процессов.
Корреляционные связи между соревновательной устойчивостью и абсолютным содержанием «древних» аминокислот у хоккеистов отсутствуют.
Таблица 3.24 сводные данные о корреляционных связях между показателями СОРЕВНОВАТЕЛЬНОЙ УСТОЙЧИВОСТИ (ПО МИЛЬМАНУ) и содержанием аминокислот у ХОККЕИСТОВ:
| Показатели | «древние» аминокислоты | серосодержащие | Произв. ГЛУ | Разветвленные аминокислоты | Циклические аминокислоты |
| АБСОЛЮТНОЕ СОДЕРЖАНИЕ АМИНОКИСЛОТ | |||||
| Сэу | |||||
| Ср | |||||
| Мэ | -ИЛЕ | ||||
| Стп | -ОРН | ||||
| ОТНОСИТЕЛЬНОЕ СОДЕРЖАНИЕ АМНИОКИСЛОТ | |||||
| Сэу | ГЛИ | ВАЛ, -ЛЕЙ | ТИР, ФЕН | ||
| Ср | |||||
| Мэ | -ИЛЕ | ||||
| Стп | |||||
| -ПРО | |||||
Если у борцов на соревновательную стабильность влияют прежде всего древние аминокислоты и циклические, то у хоккеистов в первую очередь стабильность связана с разветвленными аминокислотами и, в меньшей степени – циклическими (Табл. 3.24). Кроме того, в отличие от борцов, на саморегуляцию метаболизм аминокислот не влияет не влияет.
Сильнее всего влияние метаболизма на соревновательную стабильность проявляется на соревновательной эмоциональной устойчивости. Положительно с этим показателем связаны относительные содержания глицина, валина, тирозинаи фенилаланина. Относительное содержание глицина у хоккеистов выше, чем у борцов и легкоатлетов, и с ростом квалификации его относительное содержание возрастает, что позволяет предположить важность его роли в поддержании соревновательной эмоциональной устойчивости. Относительное содержание валина у хоккеистов поддерживает соревновательную эмоциональную устойчивость, в отличие от лейцина. Относительное содержание лейцина отрицательно связано с эмоциональной устойчивостью. Лейцина у хоккеистов меньше, чем у борцов и легкоатлетов.
3.4.4. Корреляционные связи показателей ригидности (по томскому опроснику Залевского- ТОРЗ) с содержанием аминокислот в плазме крови спортсменов
3.4.4.1 БОРЦЫ
Таблица 3.25 сводные данные о корреляционных связях между показателями ТОРЗ (ПО ЗАЛЕВСКОМУ) и содержанием аминокислот у БОРЦОВ:
| Абсолютное содержание аминокислот | «Древние» аминокислоты | Серосодержащие | Произв. ГЛУ | Разветвленные аминокислоты | Циклические аминокислоты |
| Скр | |||||
| Ар | |||||
| Пмр | |||||
| Рсо | |||||
| Ср | -ТАУ | ||||
| Ур | |||||
| ОТНОСИТЕЛЬНОЕ СОДЕРЖАНИЕ АМИНОКИСЛОТ | |||||
| Скр | |||||
| Ар | |||||
| Пмр | |||||
| Рсо | -СЕР | -ГЛН, | ВАЛ, ИЛЕ, ЛЕЙ | ФЕН | |
| Ср | -ГЛН, АРГ | ФЕН | |||
| Ур | |||||
| Всего 20 | 2 | 1 | 7 | 5 | 5 |
Высокие показатели содержания в крови серина и глутамина есть показатели недостаточности метаболизма, и борец ищет возможности чтобы избавиться от них поиском новых приемов, действий, движений.
Все незаменимые аминокислоты (вал, лей иле и фен) участвуют в глюконеогенезе, что связано с глюкокортикоидами. Борцу излишняя глюкокортикоидная функция не нужна. Это маркеры катаболиченских процессов, и чрезмерная стрессовая нагрузка делает борца осторожнее.
Повышенные количества глутамина в крови свидетельствуют о недостаточно мощных анаболических и детоксикационных механизмах в печени и почнах. И у борцов он снижает неофобию. А аргинин – протектор для нейрона [22,24,158].
Фенилаланин – предшественник адреналовых и тиреоидных гормонов, его избыток – мало гормонов и возрастает неофобия.
Показатели ригидности ТОРЗ дают понятие о склонности к формированию фиксированных форм поведения (Табл.3.25). Автор методики определяет это как субклиническое проявление нарушений функций мозга. Собственно, накопление ФФП в онтогенезе вполне естественно, и с возрастом это явление проявляет себя просто потому, что фиксированным формам событий в окружающей среде соответствуют уже проверенные ФФП, и, тем не менее, утрачивая опыт поиска для себя новых задач головной мозг утрачивает свою основную функцию – формирование новых решений.
У борцов из всех видов ФФП с метаболизмом аминокислот связаны только ригидность как состояние (РСО) и сенситивная ригидность (СР). Это показатели склонности формировать ФФП в экстремальных ситуациях (РСО), и в новых условиях – неофобия (СР).
РСО связан отрицательно с серином и глутамином. То есть способность находить решения в экстремальных условиях тем выше, чем выше в крови относительное содержание серина и глутамина. Разветвленные аминокислоты (валин, лейцин, изолейцин) и фенилаланин своим повышенным содержанием снижают способность находить новые решения в экстремальных условиях.
Если РСО и СР нарастают, значит борец утрачивает возможность поиска новых решений в экстремальных ситуациях (РСО) и боится нового, то есть утрачивает способность рисковать, используя новые решения (СР). Метаболизм аминокислот не влияют на симптомокомплекс ригидности, ни на актуальную ригидность, ни на преморбидную ригидность ни на установочную, только на показатели, определяющие стремление к поиску новых решений и готовность их находить и использовать. Таурин, серин и глутамин связаны с показателями РСО и СР отрицательной сильной связью, и их повышенное содержание в крови помогает борцам быть смелее в поиске путей к победе. А аргинин, фенилаланин и разветвленные аминокислоты, точнее их относительное содержание в крови у борцов способствует использованию уже готовых решений в новых условиях и в экстремальных условиях.
У борцов связи между аминокислотным составом крови и склонностью к фиксированным формам поведения отмечены только в отношении РСО (ригидность в экстремальных ситуациях) и СР (неофобия). Относительное содержание аминокислот имеет 9 связей с показателями ТОРЗ, и только одна связь абсолютного содержания таурина и СР (отрицательная, чем выше содержание таурина, тем ниже неофобия). Кроме того, СР связана с относительным содержанием глутамина (-) аргинина (+) и фенилаланина (+). Ригидность в экстермальных состояниях (РСО) имеет отрицательные связи с относительным содержанием серина и глутамина, и положительные связи с отностельным содержанием всех разветвленных аминокислот и фенилаланином.
3.4.4.2 ХОККЕИСТЫ
Таблица 3.26 сводные данные о корреляционных связях между показателями ТОРЗ (ПО ЗАЛЕВСКОМУ) и содержанием аминокислот у ХОККЕИСТОВ:
| Абсолютное содержание аминокислот | «древние» аминокислоты | серосодержащие | Произв. ГЛУ | Разветвленные аминокислоты | Циклические аминокислоты |
| СКР | |||||
| АР | |||||
| ПМР | |||||
| РСО | |||||
| СР | |||||
| УР | |||||
| Относительное содержание аминокислот | |||||
| СКР | |||||
| АР | |||||
| ПМР | -ГЛИ | ЛЕЙ | |||
| РСО | |||||
| СР | АРГ | ||||
| УР | -ГЛИ | ||||
Корреляционных связей между показателями ТОРЗ и абсолютным количеством аминокислот у хоккеистов нет.
Прежде всего следует отметить, что у хоккеистов глутаминовая кислота (ее абсолютное количество) снижает ригидность как свойство личности. За счет того, что глутаминовая кислота является возбуждающим медиатором нервной системы. Антагонистом в этом деле у хоккеистов является относительное содержание цистеина. Что вполне логично, поскольку серосодержащие аминокислоты являются тормозными медиаторами, цистеин дает начало другим серосодержащим аминокислотам, обладающим мощным тормозным эффектам, например, таурину (Табл. 3.26).
Аргинин – нейропротектор [22,24,158]. Скорее всего предшествующие травмы, оставляющие в памяти след, способствуют повышению его количества в ткани мозга,
Пролина у легкоатлетов в разы больше (абсолютно и относительно) чем у представителей других видов спорта. Пролин – маркер легкоатлетов. К занатиям легкой атлетикой склонны лица с высоким содержанием пролина в крови. С ростом мастерства его количество снижается. При этом делаются более прочными суставы и связки. Соединительная ткань меняется под влиянием тренировок, параллельно увеличивается содержание глутаминовой кислоты, а содержание глутамина падает. Уровень пролина у легкоатлетов отрицательно связан с нейротизмом, то есть положительно повышает активность левого полушария. Падение уровня пролина с ростом мастерства способствует повышению (относительному) активности правого полушария, то есть усиливает (косвенно) экстраверсию. Утрачивая что-то от «мозгового» типа, легкоатлеты приобретают с ростом квалификации «печеночные» свойства. В результате МСМК – сверхмозговики с элементами печеночности.
РЕЗЮМЕ ПО 3 ГЛАВЕ
На основании корреляционного анализа динамики количества аминокислот в плазме крови спортсменов различных спецификаций с параметрами их морфологических, физиологических и психологических особенностей, то есть их важнейших адаптивных систем, совершенствующихся в процессе тренировочного процесса, было установлены следующтие зависимости.
МОРФОЛОГИЧЕСКИЕ ПОКАЗАТЕЛИ
В совершенствование морфолгических структур организма спортсмена в разных видах спорта можно выделить «неспецифические» и «специфические регуляторы-аминокислоты. Так у всех спортсменов в регуляции их морфологических структур принимают участие такие аминокислоты как аспартат, глицин, гистидин, аргинин. Этот порядок аминокислот приведен по мере уменьшения корреляционных связей с морфометрическими показателями. Так, например, аспарагиновая кислота имеет у борцов 15 корреляционных связей (максимальное колтчество из всех групп), а аргинин только 1.
Однако имеются и специфические аминокислоты- регуляторы. Так у легкоатлетов в регуляции морфологических показателей большое значениеи имеют аминокислоты, имеющие в своем составе оксигруппы- треонин и серин (11 и 6 корреляционных связей), а также аланин и глутамин – по 7 корреляционных связей.
У борцов такими специфическими регуляторами являются цистеин (7 корр. связей) и глутамат (3 корр. связи). У хоккеистов- метионин, таурин и глутамат- по 5 корреляционных связей.
Больше всего аминокислот принимает участие в регуляции морфологических особенностей спортсмена у легкоатлетов-11 и меньше всего у борцов-7.
ФИЗИОЛОГИЧЕСКИЕ ПОКАЗАТЕЛИ КАРДИОРЕСПИРАТОРНОЙ СИСТЕМЫ
По физиологическим показателям к «неспецифическим» регуляторам можно отнести аспартат, лизин, фенилаланин.
При максимальном числе корреляционных связей по аспартату у борцов- 4. Обращает на себя внимание то обстоятельство, что у борцов в регуляции их кардиореспираторной системы принимает участие большинство исследуемых аминокислот -13. Среди которых по 3-4-5 корреляционных связей имеют такие как аспартат, треонин, орнитин, цистеин, пролин, метионин, лейцин, а остальные аминокислоты по 1-2 корреляционной связи.
К специфическим аминокислотам в регуляции физиологических возможностей легкоатлетовможно отнести всего 4 аминокислоты, среди которых доминирует триптофан-5 корреляционных связей. Что видимо и определяет отмеченные нами выше резкие изменения его количества при росте мастерства легкоатлетов. Среди других амирнокислот- аспартат, метионин и лизин, имеющие по 2 корреляционные связи.
У хоккеистов ведущее значение в регуляции их кардиореспираторной системы имеют глутамат, глутамин, аргинин, пролин и глицин- по 4-5 корреляционных связей. Как видим это в основном метаболиты глутаминовой кислоты. Содержание перечисленных метаболитов существенно изменяетсяри росте мастерситвва хоккеистов.
ПСИХОЛОГИЧЕСКИЕ ОСОБЕННОСТИ
В регуляции психолгических особенностей спортсменов всех групп принимает примерно равном количестве аминокислот. У борцов и легкоатлетов по 11 аминоркисот, у хоккеистов-8.
Такие аминокислоты как глицин, аланин, валин, лейцин, изолейцин, фенилаланин, тирозин являются неспецифическими и принимают участие в регуляции психологических особенностей у всех групп спортсменов. Отмечается по 1-2-3 корреляционные связи с этими аминокислотами.
Для борцов ведущее значение в регуляции психолгических особенностей имеет глутамат-4 корреляционные связи, несколько меньшее глутамин и триптофан-по 3 корреляционные связи.
Для легкоатлетов в этом плане большее значение имеет аргинин и пролин – 5 и 4 корреляционные связи соответственно. У хоккеистов ведуще значение в регуляции их психики имеет таурин-3 корреляционные связи.
Таким бразом в регуляции адаптивных систем борцов преобладают дикарбоновые аминокислоты- аспартат и глутамат- образующиеся в больших количествах в больших количествах в мышцах и мозге.
У хоккеистов – таурин, глутаминовая кислота и ее производные- метаболиты ситезируемые преимущественно в печени
У легкоатлетов- аспартат, аланин, глицин, пролин, тирозин, триптофан, аргинин, серин, треонин- то есть амнокислоты использующиеся в большей степени в метаболических процессах мозга, нервной и соединительной ткани. То есть адаптивные системы лекоатлетов имеют гораздо более сложную аминокислотную регуляцию.
ГЛАВА 4. ПАСПОРТА ОТДЕЛЬНЫХ АМИНОКИСЛОТ, НАИБОЛЕЕ ШИРОКО ПРИМЕНЯЕМЫХ В СПОРТЕ И ФИТНЕСЕ, СОСТАВЛЕННЫЕ ПО МАТЕРИАЛАМ НАШИХ ИССЛЕДОВАНИЙ
На наш взгляд наибольшую ценность для целенаправленного применения аминокислот будет представлять сопоставление изменений количеств аминокислот как по абсолютным, так и по относительным показателям с динамикой морфологических, физиологических и психологических параметров у спортсменов различных спецификаций и различного уровня мастерства.
ТАУРИН
4.1 Графическое изображение количества таурина в плазме крови спортсменов разных видов спорта и в зависимости от уровня квалификации.
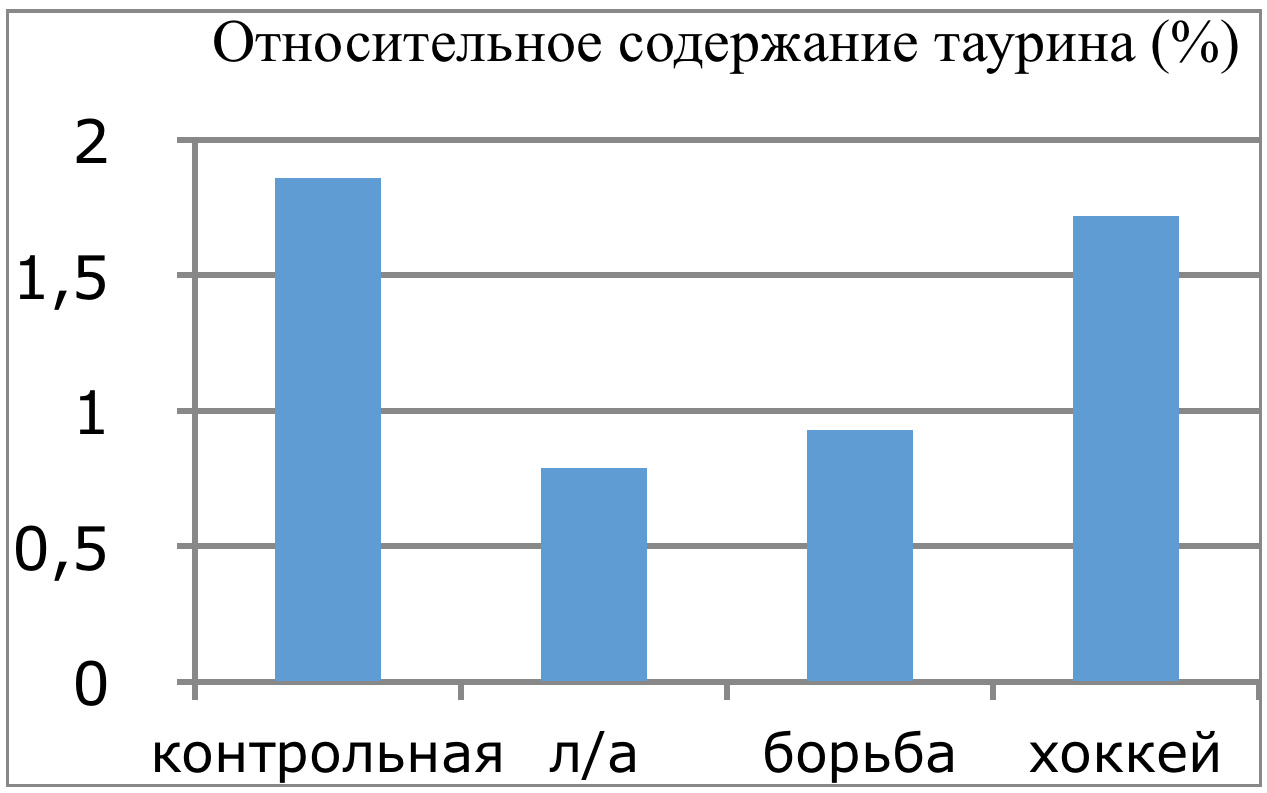
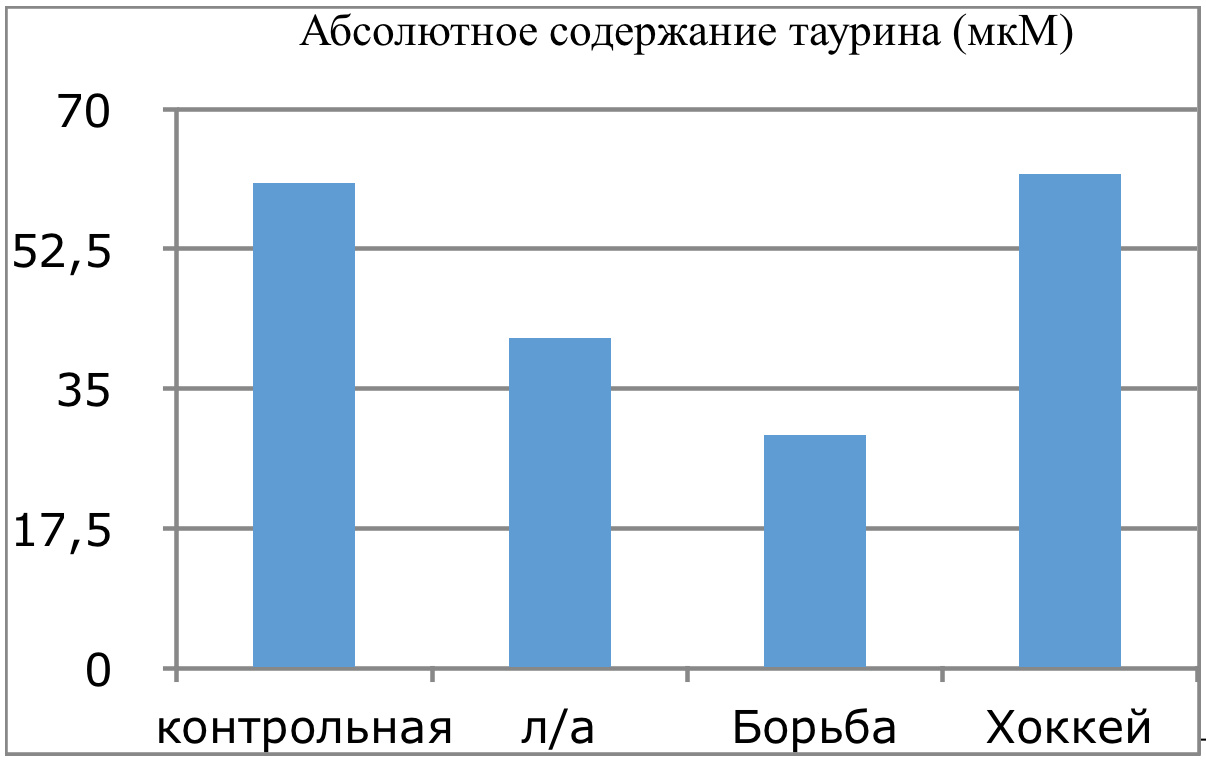
Абсолютное содержание таурина (мкМ) в зависимости от квалификации:
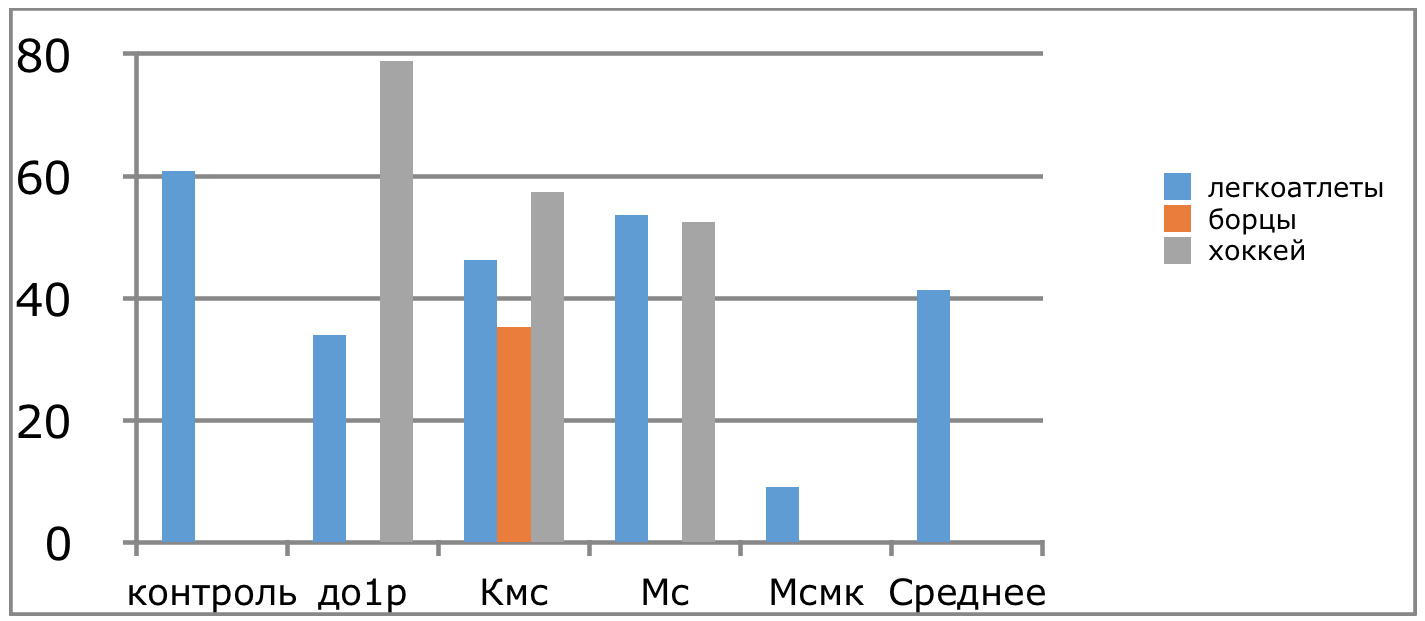
Относительное содержание таурина (%) в зависимости от квалификации:
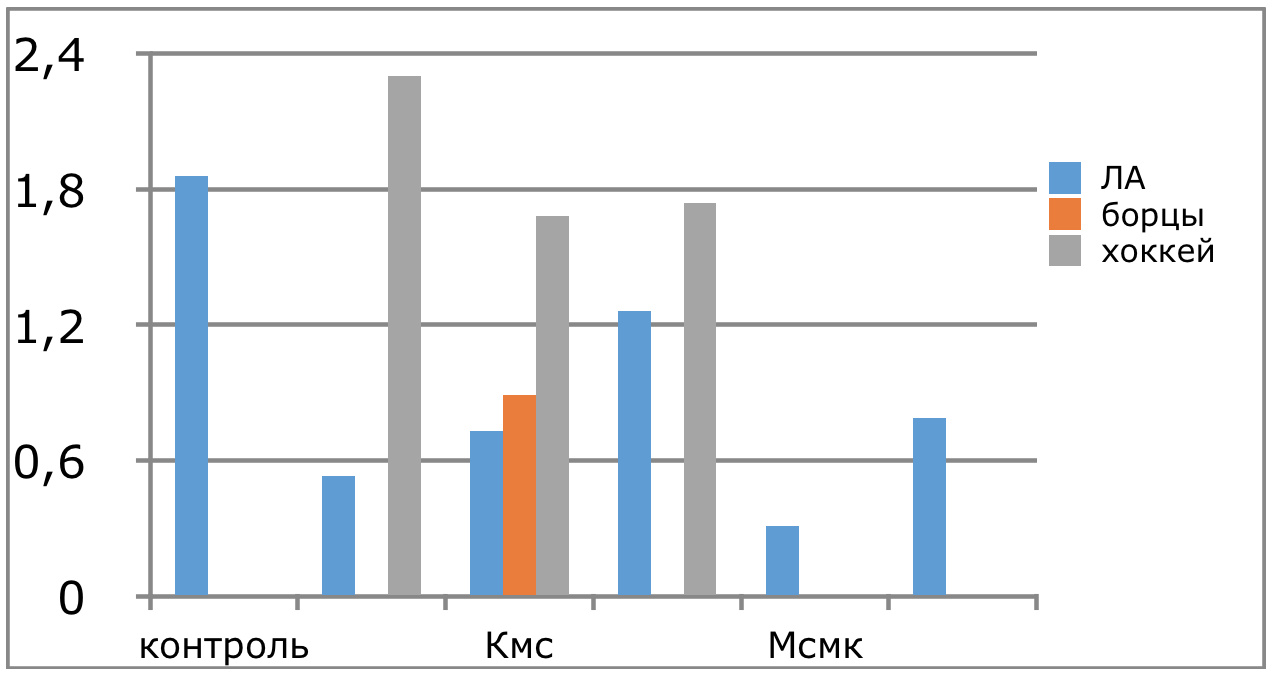
Таблица 4.1 Корреляционные связи у борцов, ЛА и хоккеистов:
| БОРЦЫ | ЛЕГКОАТЛЕТЫ | ХОККЕИСТЫ | ||||
| Показатели | Тау (относ. к-во) | Тау (абсолют. к-во) | Тау (относит.к-во) | Тау (абсолютное количество) | Тау (относит. к-во) | Тау (абсолют,к-во) |
| отнширпл | -,742* | |||||
| отндлрук | -,805* | -,798* | ||||
| Отнжира | ,899* | |||||
| Перзад | ,817* | ,799* | ||||
| Кум | -,999* | |||||
| экстраверт | -,976* | |||||
| сансамоч | -,980* | |||||
| Санактив | -,978* | |||||
| Торзсенс | -,833* | + | + | |||
| @__S_ | -,928* | -,890* | ||||
| ** Корреляция значима на уровне 0.01 (2-сторон.). | ||||||
| * Корреляция значима на уровне 0.05 (2-сторон.). | ||||||
ОБЩЕЕ КОЛИЧЕСТВО:
| таурин | борцы | ЛА | хоккеисты | |||
| коррел | В т.ч. отр | Коррел | В т.ч. отр | коррел | В т.ч. отр. | |
| Морфметрия | 1 | 1 | 1 | 5 | 3 | |
| Кардиореспират. с-ма | 2 | 2 | ||||
| Психолочические особ. | 1 | 1 | 3 | 3 | ||
| Всего 13 | 2 | 2 | 3 | 2 | 8 | 6 |
Содержание таурина (абсолютное и относительное) у легкоатлетов и у борцов достоверно ниже, чем в контрольной группе, а у хоккеистов не отличается от контроля. По мере возрастания квалификации у легкоатлетов содержание таурина у легкоатлетов возрастает почти до уровня контроля у мастеров спорта, с падением почти на порядок по отношению к контролю у МСМК (Графики 4.1).
У борцов и хоккеистов уровень таурина в плазме крови снижается, что особенно видно по абсолютному показателю(Табл.4.1).
Корреляционные связи таурина у легкоатлетов наблюдаются с относительной жирностью (положительная связь) и с тонусом симпатической нервной системы (отрицательная). С большим количеством таурина легкоатлеты имеют больше жира и спокойствия, это скорее марафонцы, чем спринтеры. Кроме того, большое количество таурина усиливает у легкоатлетов неофобию.
У борцов содержание таурина тоже связано с неофобией, но отрицательно и отрицательно же с относительной шириной плеч. То есть таурин осуществляет у борцов метаболическую связь между широкоплечестью и способностью быстро находить новые решения в новых условиях.
У хоккеистов таурин метаболически связывает короткие руки с интраверсией. Кроме того, таурин снижает самочувствие и активность по самооценке, и понижает критическую удельную мощность при тестировании работоспособности. Таким образом, содержание таурина пронизывает сущность хоккеиста, связывая воедино морфологические, физиологические функциональные и психологические параметры – темперамент и самооценку.
АСПАРАГИНОВАЯ КИСЛОТА
Аспарагиновая кислота относится к заменимым аминокислотам. Известна как нейропротектор.
Повышает работоспособность, ускоряет восстановление при утомлении. В нашем исследовании наибольшее количество аспарагиновой кислоты содержится в крови у легкоатлетов, как в абсолютном количестве, так и относительно, больше чем в контроле на 30% относительно и почти в 3 раза абсолютно. У борцов и хоккеистов аспарагиновая кислота в плазме крови содержится в меньшем количестве, чем в контроле. Занятия спортом по-разному влияют на содержание аспарагиновой кислоты в плазме крови спортсменов. У борцов и хоккеистов содержание аспарагиновой кислоты в крови с ростом мастерства возрастает (Графики 4.2). У легкоатлетов и так высокое содержание в крови аспарагиновой кислоты, но оно еще возрастает до уровня мастера спорта, на уровне мастера спорта международного класса ее уровень падает вдвое и более — ниже уровня контроля.
4.2 Графическое изображение количества аспарагиновой кислоты в плазме крови спортсменов разных видов спорта и в зависимости от уровня квалификации.
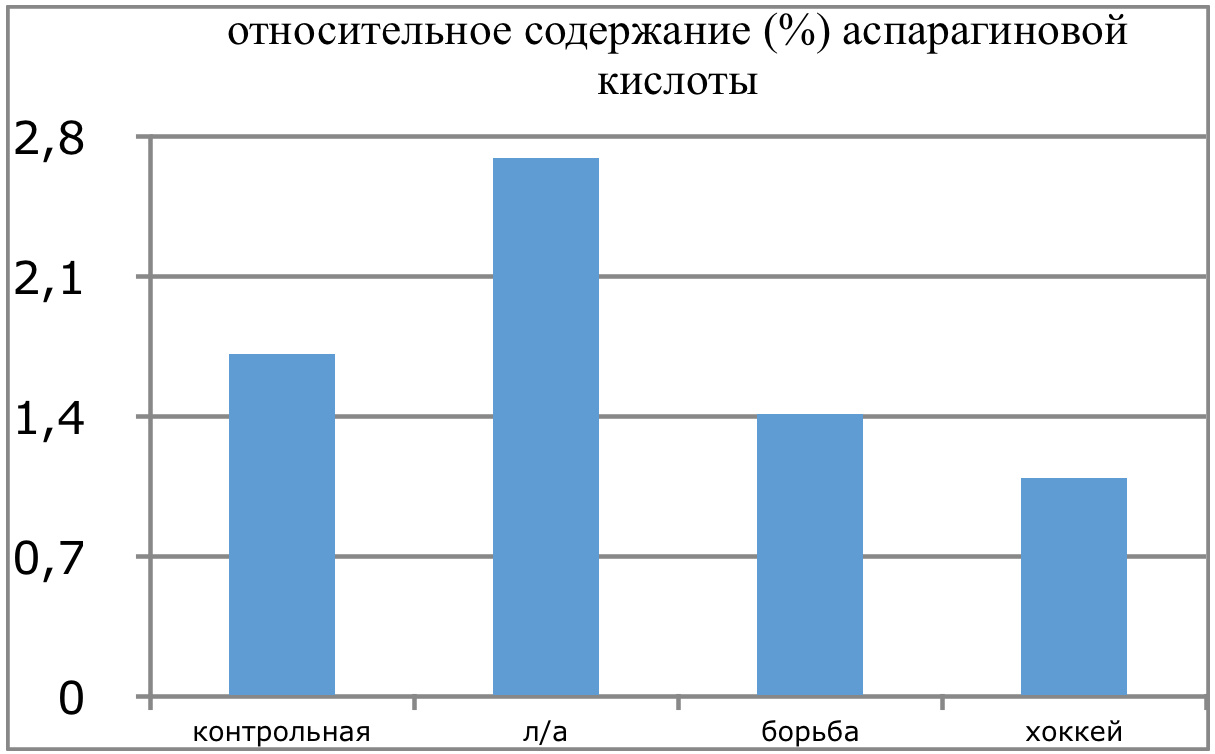
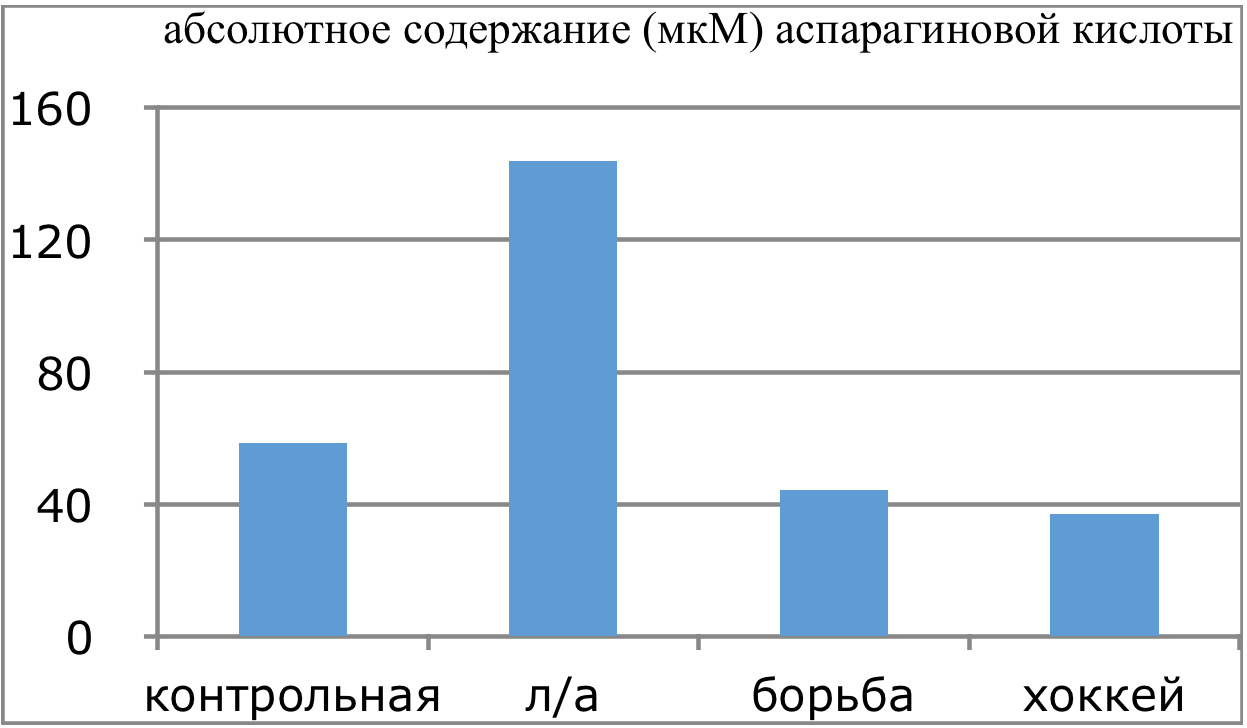
Абсолютное содержание аспарагиновой кислоты в зависимости от квалификации:
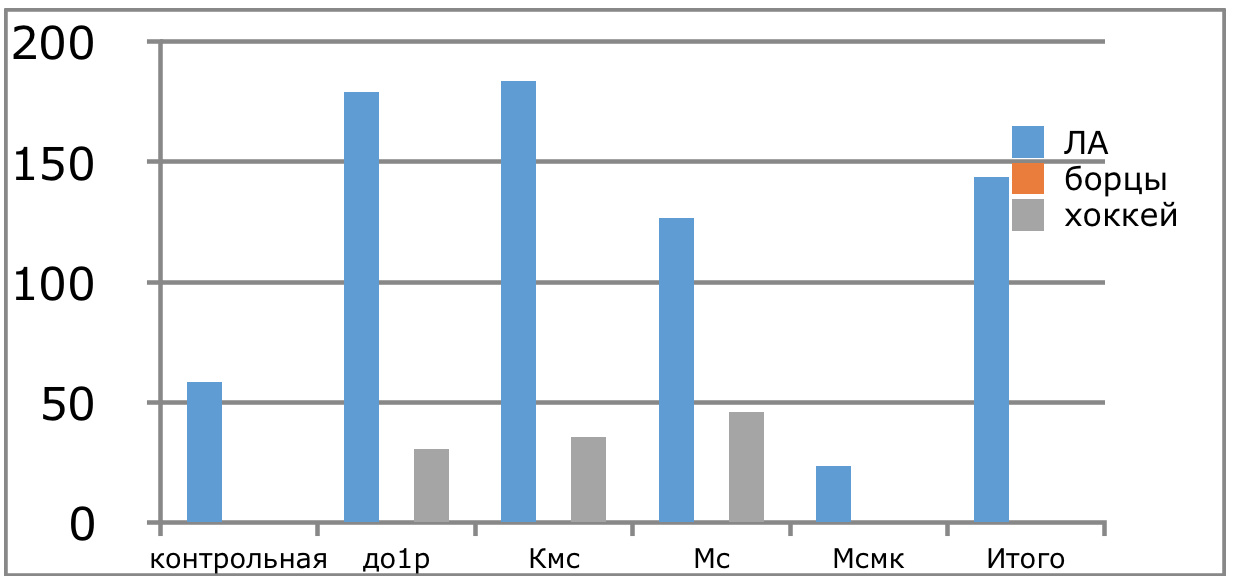
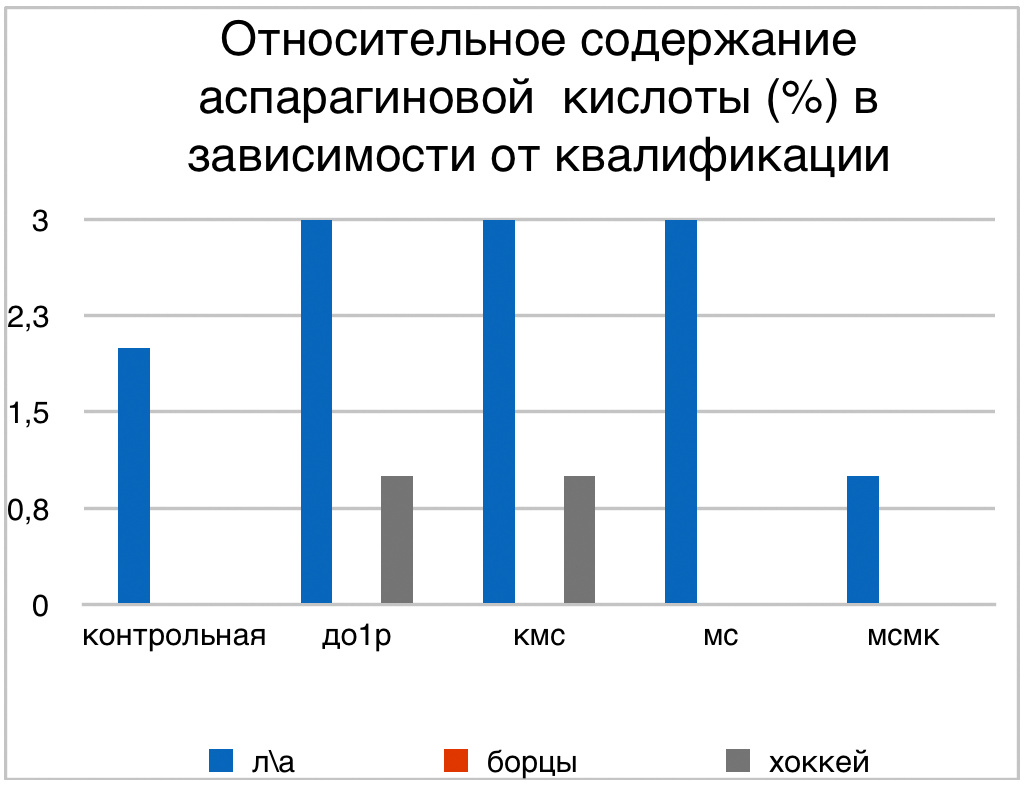
Таблица 4.2 Корреляционные связи у борцов, ЛА и хоккеистов:
| БОРЦЫ | ЛЕГКОАТЛЕТЫ | ХОККЕИСТЫ | ||||
| показатели | Асп(относит. к-во) | Асп(абсолют. к-во) | Асп(относит. к-во) | Асп(абсолют. к-во) | Асп(относит. к-во) | Асп(абсолют. к-во) |
| возраст | ,884** | ,786** | ||||
| рост | ,712* | |||||
| весор | ,682* | ,720* | ||||
| ип | ,749* | -,837* | ||||
| отнширпл | -,847* | |||||
| ширтаза | ,723* | |||||
| длруки | ,691* | |||||
| длног | ,755* | ,893** | ,854* | |||
| отндлног | ,976** | ,977** | ||||
| весжира | ,866** | ,742* | ||||
| отнжира | ,723* | |||||
| весмышц | ,678* | -,804* | -,748* | |||
| отнмышц | -,808* | |||||
| отнкостей | -,852** | -,668* | ,972** | ,957* | ||
| окрголов | -,759* | |||||
| пвцудель | ,707* | ,747* | 0,947* | |||
| ToeA | ,884* | |||||
| NISP | ,826** | ,632* | ||||
| EF_ | -,884* | |||||
| ** Корреляция значима на уровне 0.01 (2-сторон.) | ||||||
| * Корреляция значима на уровне 0.05 (2-сторон.). | ||||||
ОБЩЕЕ КОЛИЧЕСТВО:
| Асп к-та | борцы | ЛА | хоккей | |||
| коррел | В т.ч. отр | Коррел | В т.ч. отр | коррел | В т.ч. отр | |
| Морфометрия | 15 | 2 | 7 | 2 | 4 | 4 |
| Кардиореспираторная с-ма | 4 | 2 | 1 | 1 | ||
| Психологические особ-сти | ||||||
| Всего 34 | 19 | 2 | 9 | 3 | 5 | 4 |
Связи содержания аспарагиновой кислоты с параметрами тела самые сильные не у легкоатлетов, а у борцов (Табл.4.2). Аспарагиновая кислота пронизывает тело борца буквально «насквозь» — содержание этого метаболита возрастает с возрастом, положительно связано с размерами тела, пропорциями тела, со всеми компонентами тела – с мышцами и жиром, а также с удельной мощностью, которую борцы развивали при тестировании ПВЦ 170. Из физиологических показателей содержание аспарагиновой кислоты положительно связано у борцов с систолическим артериальным давлением. Единственная отрицательная связь содержания аспарагиновой кислоты у борцов – с относительным содержанием костей в теле борцов.
У легкоатлетов избыток аспарагиновой кислоты приводит к астенизации – отрицательная связь с шириной плеч и индексом пропорциональности, и положительная связь с длиной ног. При этом положительная связь с костистостью тела легкоатлета сочетается с отрицательной связью с эффективностью сердечных сокращений (фракция выброса).
У хоккеистов отмечена отрицательная связь содержания аспарагиновой кислоты в крови с абсолютной и относительной мышечной массой (противоположная по знаку зависимость у борцов). Но на тестирование работоспособности у хоккеистов содержание аспарагиновой кислоты воздействует аналогично таковому у борцов – сильная положительная связь.
Таким образом, содержание аспарагиновой кислоты в крови спортсменов зависит от вида спорта, от квалификации, и оказывает разнонаправленное влияние на морфологию и физиологию тел спортсменов в зависимости от вида спорта.
В процессе совершенствования аспарагиновая кислота наибольшее значение имеет для легкоатлетов. При том, что у МСМК ее содержание снижается, тем не менее ее уровень остается высоким по отношению к представителям других видов спорта. А у борцов имеется тенденция к росту с возрастанием мастерства. То есть в становлении борца дикарбоновые кислоты имеют ключевое значение, но если уровень глутаминовой кислоты изначально высок, то уровень аспарагиновой аминокислоты возрастает. Эти аминокислоты являются основой формирования борца.
При анализе корреляционных связей с установлено, что главным образом аспарагиновая кислота влияет на формирование структурного следа адаптации – с морфологией у борцов 20 связей, у ЛА – 7 связей, у хоккеистов – 4 связи. По 2 связи с физиологическими параметрами. На психологию влияния нет. Из чего можно сделать вывод, что давать аспарагиновую кислоту нужно только на стадии формирования спортсмена, на уровне 1 разряда, КМС, когда формируется структурный след адаптации. У мастеров и МСМК она может оказать вредное влияние.
СЕРИН
4.3 Графическое изображение количества серина в плазме крови спортсменов разных видов спорта и в зависимости от уровня квалификации:
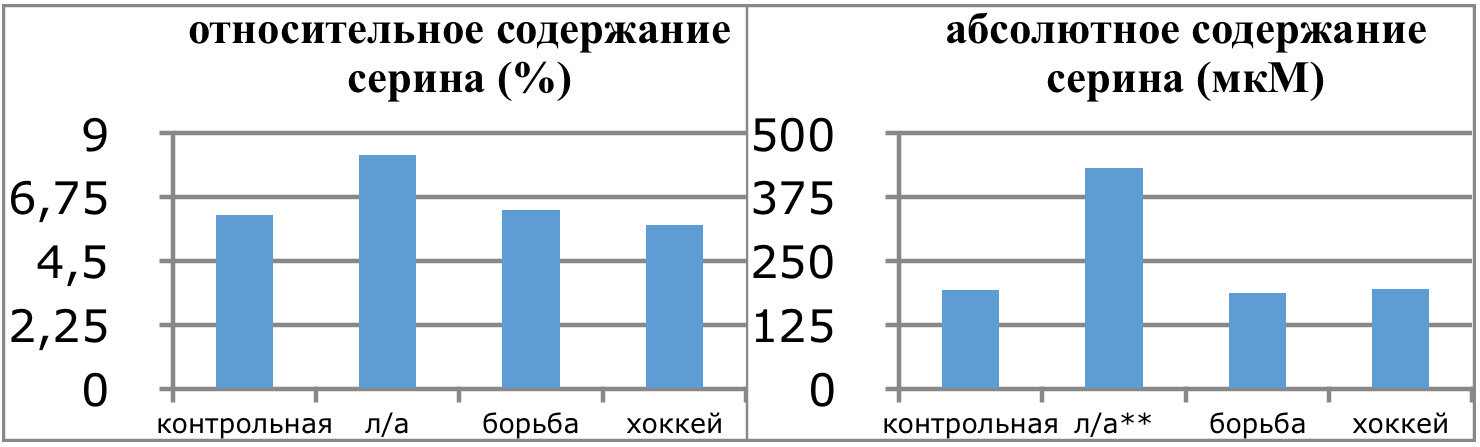
Серин (%):
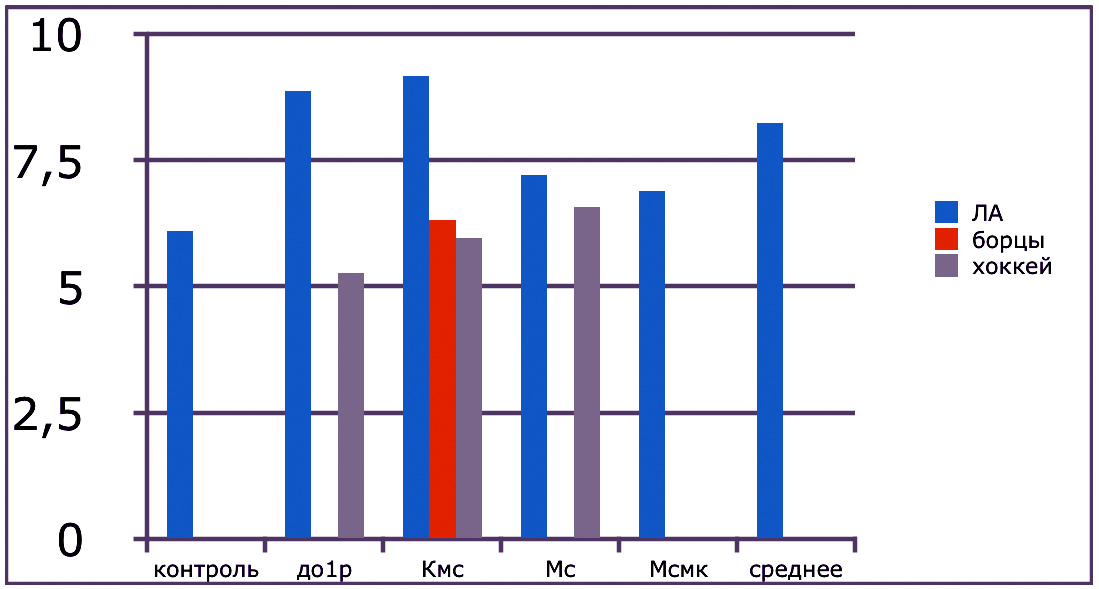
Серин (мкМ):
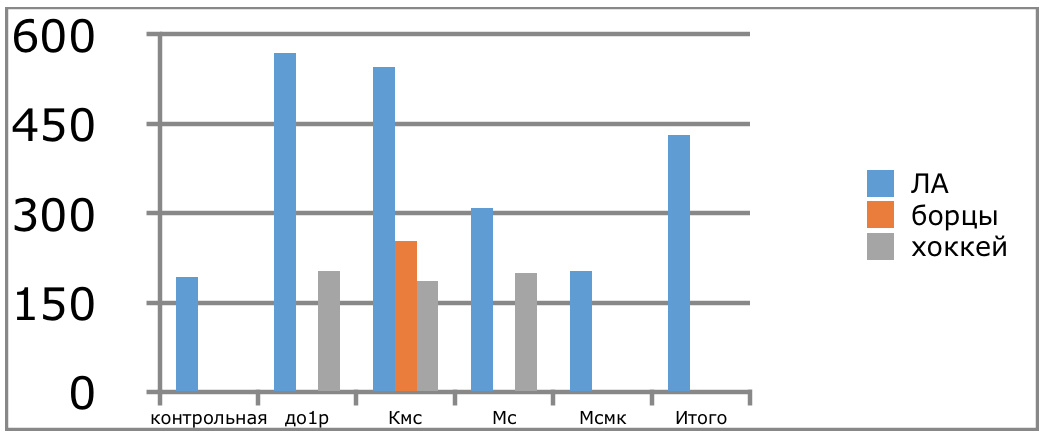
Содержание серина у легкоатлетов выше, чем в контроле и у борцов и хоккеистов, особенно это заметно (Р<0,01) в абсолютном содержании серина. С ростом квалификации у легкоатлетов и борцов содержание серина снижается, что тоже резче выражено в абсолюном содержании. У хоккеистов изменения абсолютного содержания серина с ростом квалификации нет, относительное же содержание серина у них с ростом квалификации имеет тенденцию к росту (Графики 4.3).
У ЛА резкий рост содержания серина – примерно в 2,5 раза до КМС, а затем снижается, у борцов тоже рост и снижение. В процессе совершенствования у борцов и ЛА серин необходим как универсальный метаболит – предшественник серосодержащих аинокислот, фосфолипидов, донатор метильных групп, анаболик для построения белков. У хоккеистов существенных изменений не выявлено, и для совершенствования значения не имеет.
Табл. 4.3 Корреляционные связи у борцов, ЛА, хоккеистов:
| БОРЦЫ | ЛЕГКОАТЛЕТЫ | ХОККЕИСТЫ | ||||
| Показатели | Сер(относит. к-во) | Сер(абсолют.к-во) | Сер(относит.к-во) | Сер(абсолют.к-во) | Сер(относит.к-во) | Сер(абсолют.к-во) |
| рост | ,823* | |||||
| ип | -,821* | |||||
| длтул | ,860* | |||||
| длног | ,887* | |||||
| отндлног | ,871* | |||||
| весорг | ,924* | |||||
| отнорг | ,753* | |||||
| сэу | ,824* | |||||
| РСО | -,888* | |||||
| PI_ | ,744* | |||||
| SpO2 | ,980** | |||||
| ToeA | ,935* | |||||
| @__S_ | -,934** | |||||
| EF_ | -,993** | |||||
| ** Корреляция значима на уровне 0.01 (2-сторон.). | ||||||
| * Корреляция значима на уровне 0.05 (2-сторон.). | ||||||
ОБЩЕЕ КОЛИЧЕСТВО:
| серин | борцы | ЛА | хоккей | |||
| коррел | В т.ч. отр | коррел | В т.ч. отр | коррел | В т.ч. отр. | |
| Морфометрия | 6 | 1 | 1 | |||
| Кардиореспираторная с-ма | 1 | 1 | 2 | 1 | ||
| Психологические особенности | ||||||
| Всего 10 | 1 | 7 | 3 |
У борцов содержание серина не коррелирует ни с морфологическими ни с функциональными показателями. Только с психологическими показателями – повышает соревновательную эмоциональную устойчивость и гибкость решений в экстремальных ситуациях, повышает настроение (Табл.4.3).
У легкоатлетов резкий рост содержания серина – примерно в 2,5 раза до КМС, а затем снижается. Абсолютное содержание серина увеличивает длиннотные размеры и улучшает микроциркуляцию, снижая эффективность сердечных сокращений. 3 корреляции с морфологией.
У хоккеистов сдвигает тонус вегетативной нервной системы в парасимпатическую сторону, усиливая процессы восстановления. Способствует аэробизации организма.
Таким образом, действие серина на организм спортсменов сильно зависит от характера нагрузки, определяемой видом спорта.
Серин необходим в процессе реабилитации в психологическом плане у борцов и в физиологическом у хоккеистов. В формировании структурного следа адаптации существенного значения не имеет.
ГЛУТАМИНОВАЯ КИСЛОТА
4.4 Графическое изображение количества глутаминовой кислоты в плазме крови спортсменов разных видов спорта и в зависимости от уровня квалификации:
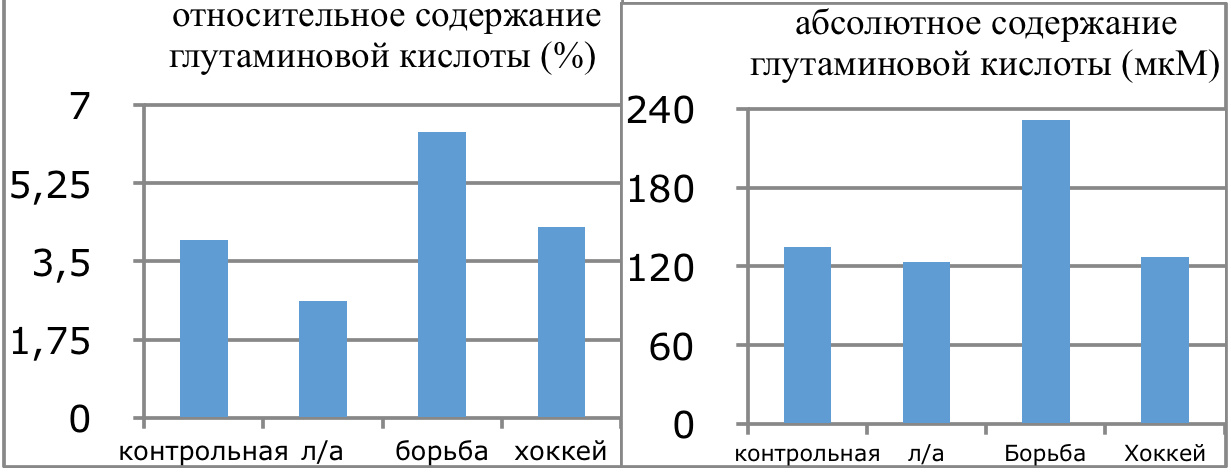
Глутаминовая кислота (%):
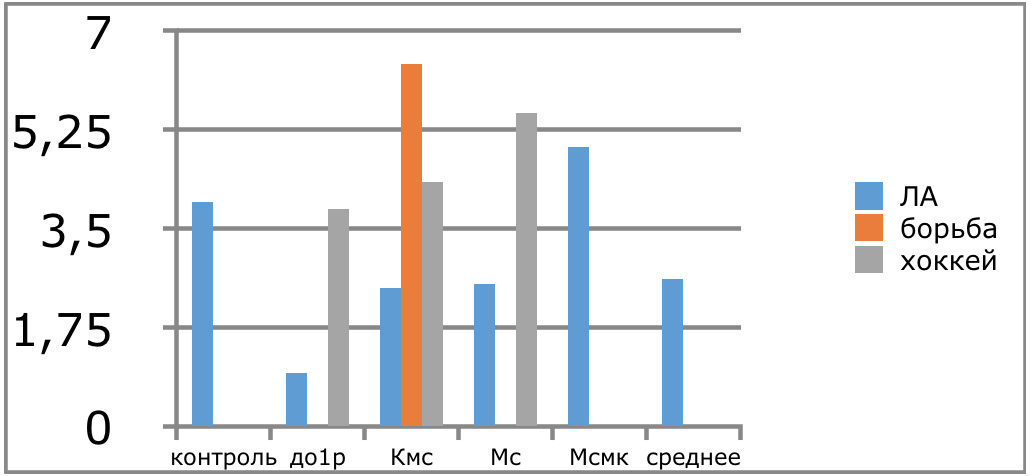
Глутаминовая кислота (Мкм):
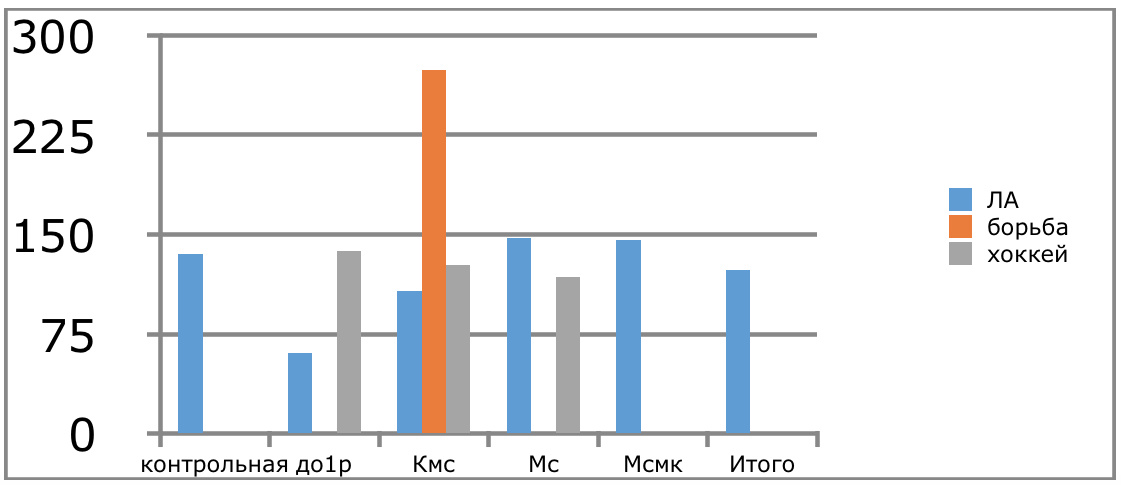
Глутаминовая кислота выделяет борцов. У борцов её больше, чем у представителей других видов спорта почти вдвое (Графики 4.4). С ростом мастерства абсолютное количество глутаминовой кислоты несколько снижается, но относительное продолжает возрастать. Содержание глутаминовой кислоты у борцов коррелирует с количеством мышечного компонента. Кроме того, глутаминовая кислота для борцов является фактором, усиливающим мотивацию, улучшает настроение. Кроме того, содержание глутаминовой кислоты в крови связано отрицательной корреляционной связью с ЦВД. Снижение ЦВД предположительно связано с тем, что высокое содержание глутаминовой кислоты повышает возбудимость нейронов центральной нервной системы и облегчает формирование первой фазы стресса (катехоламиновой) а в этой фазе повышен тонус мышц головы и шеи и мышц, обеспечивающих вдох.
Это свойство – облегчение формирования катехоламиновой фазы общего адаптационного синдрома за счет глутаминовой кислоты (отрицательная связь с ЦВД) – выделено и у легкоатлетов. Содержание глутаминовой кислоты в крови у легкоатлетов ниже, чем в контроле, но по мере роста мастерства и с возрастом количество глутаминовой кислоты у легкоатлетов возрастает.
Таблица 4. 4 Корреляционные связи у борцов, ЛА, хоккеистов:
| БОРЦЫ | ЛЕГКОАТЛЕТЫ | ХОККЕИСТЫ | ||||
| Показатели | Глу (относит. К-во) | Глу(абсолют.к-во) | Глу(относит.к-во) | Глу(абсолют.к-во) | Глу(относит.к-во) | Глу(абсолют.к-во) |
| возраст | ,840* | |||||
| ширпл | ,709* | |||||
| ширтаза | ,845** | |||||
| Отнширтаз | ,763* | |||||
| отнмышц | ,836** | ,764* | -,754* | -,798* | ||
| отнорг | -,727* | |||||
| саннастр | ,809* | |||||
| ср | -,770* | |||||
| мэ | ,730* | ,870** | ||||
| пмр | -,957* | |||||
| PI_ | ,644* | |||||
| SV_ | -,787** | -,943** | ||||
| Fw_ | -,745* | -,868** | -,919* | |||
| CO_ | -,911* | |||||
| CI_ | -,900* | |||||
| DO2 | -,909* | |||||
| ** Корреляция значима на уровне 0.01 (2-сторон.). | ||||||
| * Корреляция значима на уровне 0.05 (2-сторон.). | ||||||
ОБЩЕЕ КОЛИЧЕСТВО:
| Глутаминовая к-та | борцы | ЛА | хоккей | |||
| коррел | В т.ч. отр | коррел | В т.ч. отр | коррел | В т.ч. | |
| Морфометрия | 3 | 1 | 1 | 5 | 2 | |
| Кардиреспираторная с-ма | 2 | 2 | 1 | 1 | 5 | 5 |
| Психологические особенности | 4 | 1 | 1 | 1 | ||
| Всего 22 | 9 | 4 | 3 | 2 | 10 | 7 |
У хоккеистов глутаминовая кислота связана с шириной плеч и таза положительной корреляционной связью, и, как ни странно, отрицательной связью с мышечным компонентом тела. Кроме того, отрицательная связь содержания глутаминовой кислоты у хоккеистов отмечается с ударным объемом, с минутным объемом крови и интенсивностью тканевого метаболизма (индекс доставки кислорода) (Табл.4.4).
Давать надо глутаминовую кислоту легкоатлетам, борцам может быть и надо давать, но они сами ее производят, прежде чем давать надо разобраться. У хоккеистов количество глутаминовой кислоты существенных изменений в крови не претерпевает, смысла давать глутамат хоккеистам очевидно никакого нет.
ГЛУТАМИН
4.5 Графическое изображение количества глутамина в плазме крови спортсменов разных видов спорта и в зависимости от уровня квалификации:
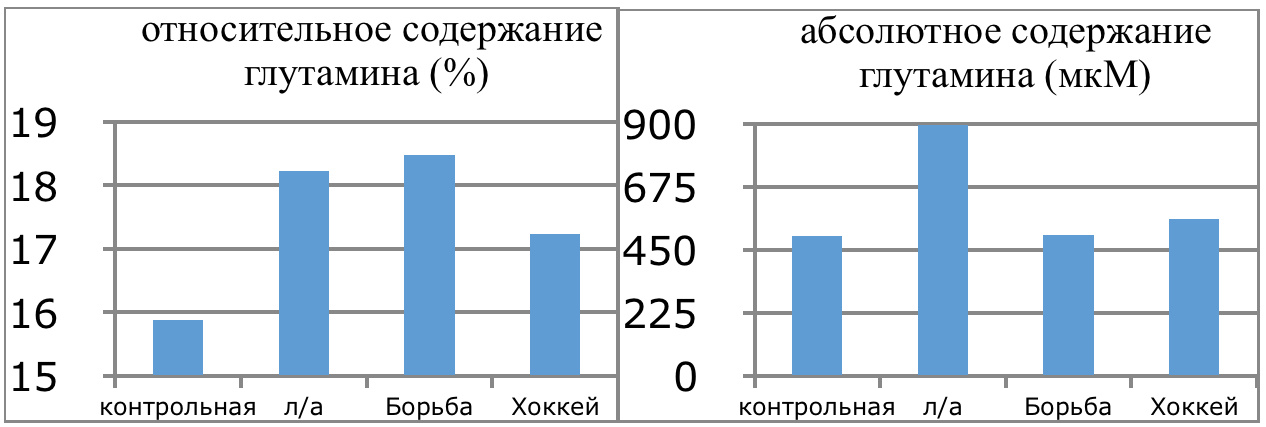
Глутамин (%):
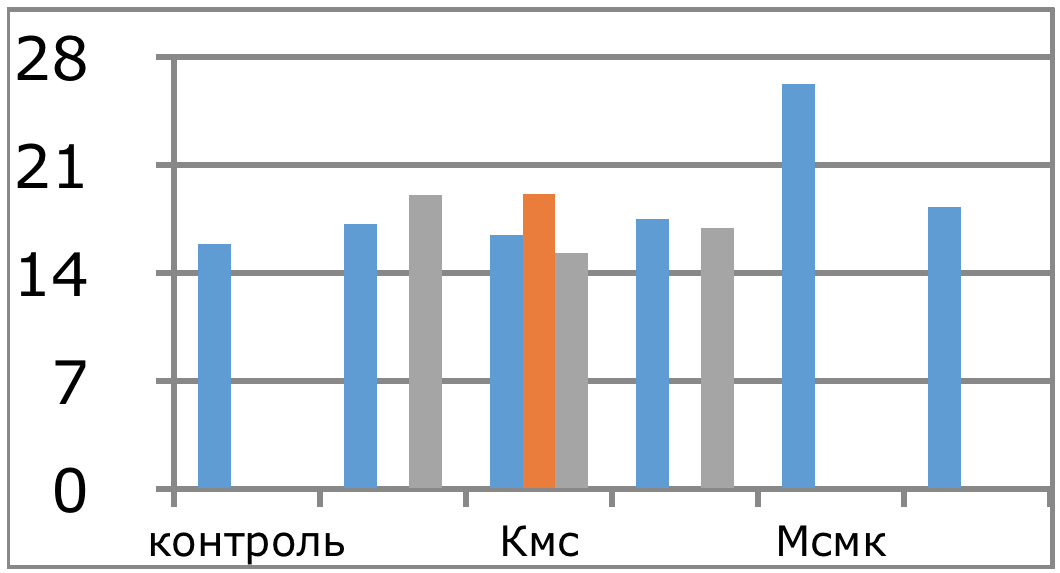
Глутамин (мкМ):
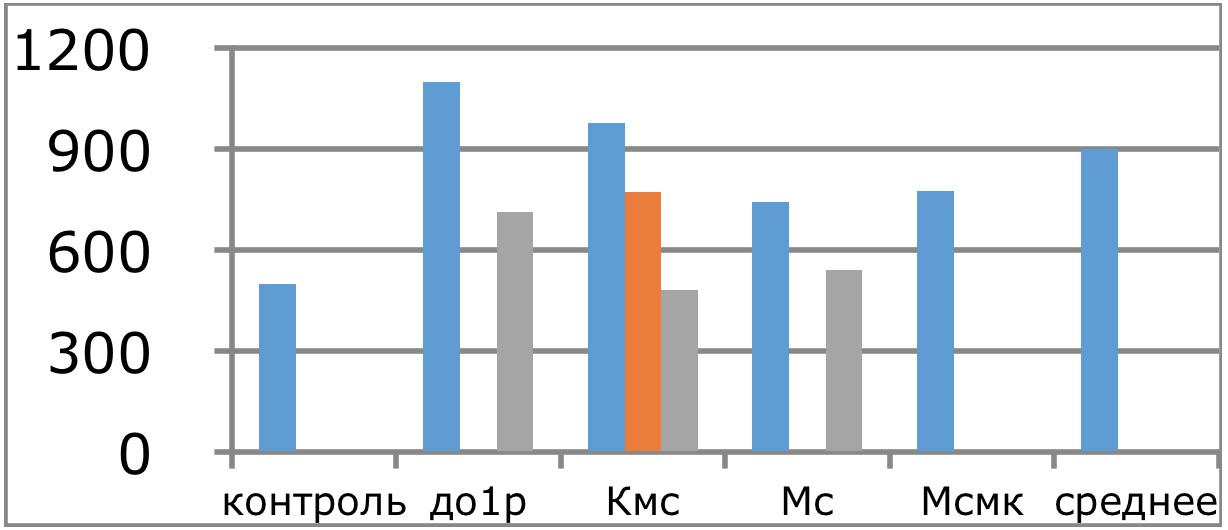
У борцов глутамин с ростом квалификации снижается (Графики 4.5). Корреляционные связи глутамина наблюдаются только с психологическими параметрами – глутамин снижает настроение и делает поведение борцов в условиях стресса более гибким.
Сущность легкоатлета пронизана глутамином насквозь. Рост, ширина таза и длина ног имеют отрицательную связь с глутамином, то есть рост и размеры нижней части тела у легкоатлетов под влиянием глутамина уменьшаются. Повышается мотивация для занятий легкой атлетикой, повышается осторожность и любопытство.
Действие глутамина на хоккеистов – чисто физиологическое – повышается критическая удельная мощность, то есть срыв дыхания у хоккеистов происходит позже, если в крови у них относительно много глутамина, да и насыщение крови кислородом выше.
Глутамин больше связан не с соматическими функциями, а с нервной системой. Его нужно давать уже сформированным спортсменам – легкоатлетам ((Табл. 4.5).
Таблица 4.5 Корреляционные связи у борцов, ЛА и хоккеистов:
| БОРЦЫ | ЛЕГКОАТЛЕТЫ | ХОККЕИСТЫ | ||||
| Показатели | Глн(относит.к-во) | Глн(абсолют. к-во) | Глн(относит. к-во) | Глн(абсолют. К-во) | Глн(относит. К-во) | Глн(абсолют. К-во) |
| рост | -,822* | |||||
| ип | ,833* | -,829* | ||||
| отндлтул | ,859* | |||||
| ширтаза | -,871* | |||||
| длног | -,852* | |||||
| весорг | ,884* | |||||
| кум | ,999* | |||||
| санактив | ,994** | |||||
| саннастр | -,772* | ,998* | ||||
| мэ | 1,000** | 1,000** | ||||
| РСО | -,937** | 1,000** | 1,000** | |||
| торзсенс | -,867* | -1,000** | -1,000** | ,977* | ||
| SpO2 | ,889* | ,924** | ||||
| ToeA | ||||||
| NISP | ||||||
| NIDP | ||||||
| @__S_ | -,915* | |||||
| ** Корреляция значима на уровне 0.01 (2-сторон.) | ||||||
| * Корреляция значима на уровне 0.05 (2-сторон.). | ||||||
ОБЩЕЕ КОЛИЧЕСТВО:
| глутамин | борцы | ЛА | хоккей | |||
| коррел | В т.ч. отр | коррел | В т.ч. отр | коррел | В т.ч отр.. | |
| Морфометрия | 7 | 4 | ||||
| Кардиореспираторная система | 4 | 1 | ||||
| Психологические особенности | 3 | 3 | 1 | |||
| Всего 15 | 3 | 3 | 7 | 4 | 5 | 2 |
ПРОЛИН
4.6 Графическое изображение количества пролина в плазме крови спортсменов разных видов спорта и в зависимости от уровня квалификации:
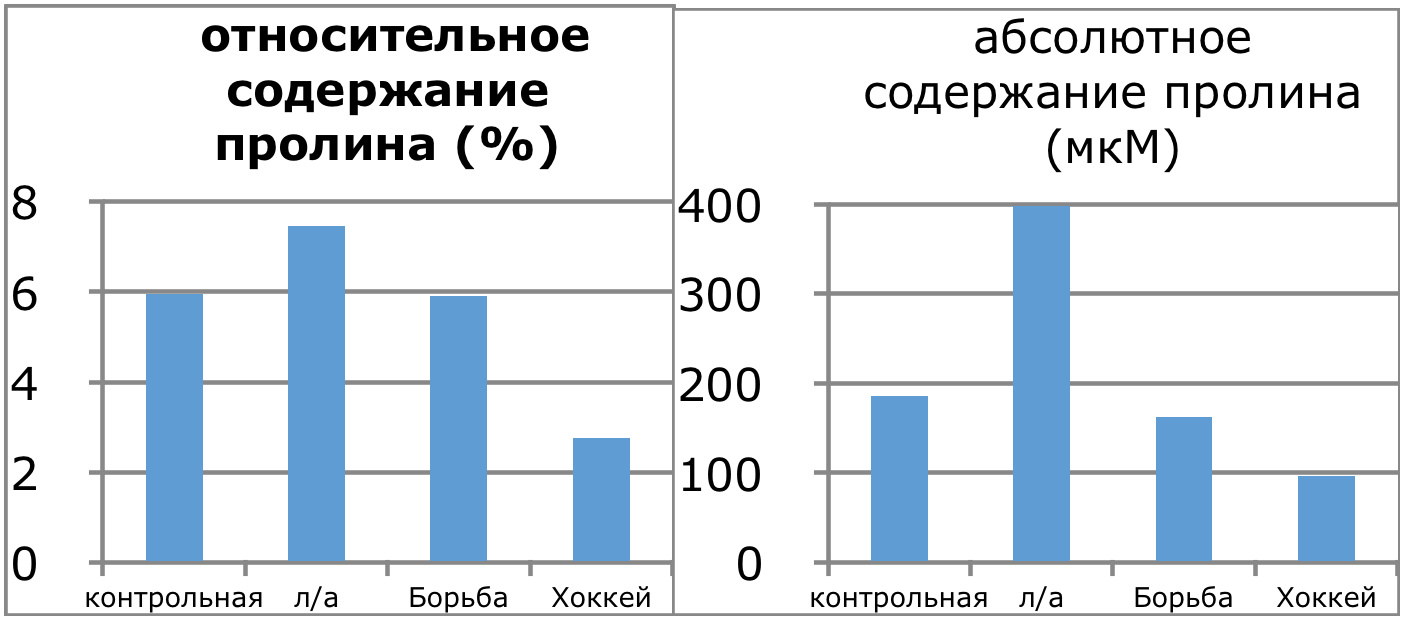
Пролин (Мкм):
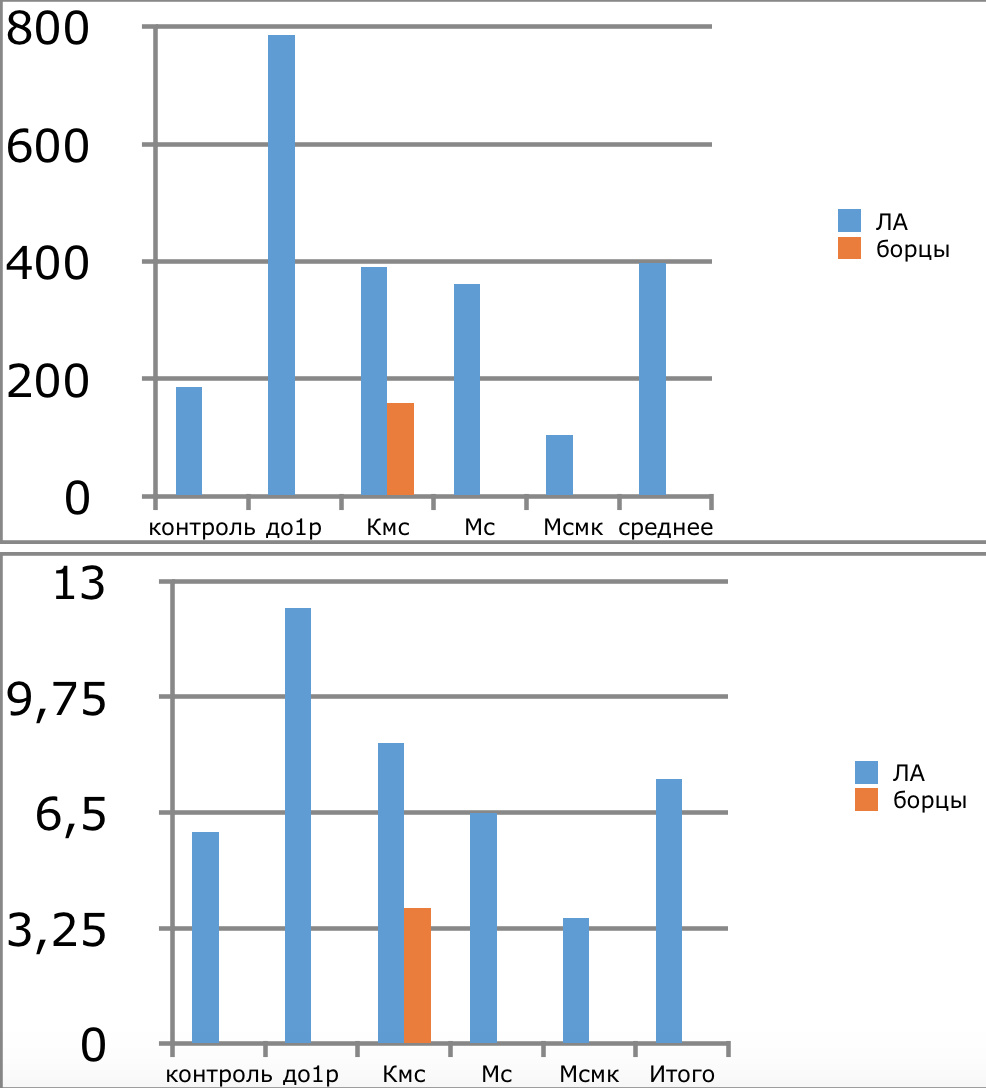
Пролин (%).
Значение пролина для борцов – влияет на размеры таза и длина ног, а еще – отрицательная связь с нейротизмом. То есть пролин увеличивает контроль левого полушария над лимбикой, что в свою очередь повышает соревновательную стабильность.
У хоккеистов пролин на стабильность влияет противоположным образом, то есть снижает её. А также содержание пролина в крови у хоккеистов имеет отрицательную корреляционную связь с шириной плеч и таза (Табл.4.6).
Важен как антистрессовое вещество у легкоатлетов, но не у борцов и отрицательно воздействует на хоккеистов. То есть в период соревнований у легкоатлетов есть смысл применять пролин.
Таблица 4. 6 Корреляционные связи у борцов, ЛА и хоккеистов (БОРЦЫ, ЛЕГКОАТЛЕТЫ, ХОККЕИСТЫ):
| Показатели | Про(относит.к-во) | Про(абсолют. К-во) | Про(относит.к-во) | Про(абсолют.к-во) | Про(относит.к-во) | Про(абсолют.к-во) |
| Отнширп | -,718* | |||||
| Ширтаз | ,681* | -,711* | -,731* | |||
| отншитаза | -,743* | -,822* | ||||
| длног | ,684* | ,692* | ||||
| отнорг | ,889* | |||||
| санактив | ,960* | |||||
| стп | 1,000** | 1,000** | -,954* | |||
| СР | ,976* | |||||
| УР | ,997** | |||||
| ** Корреляци значима на уровне 0.01 (2-сторон.). | ||||||
| * Корреляция значима на уровне 0.05 (2-сторон.). | ||||||
ОБЩЕЕ КОЛИЧЕСТВО:
| пролин | борцы | ЛА | хоккей | |||
| коррел | В т.ч. отр | коррел | В т.ч. отр | коррел | В т.ч. | |
| Морфметрия | 3 | 1 | 5 | 5 | ||
| Кардиореспираторная система | ||||||
| Психологические особености | 4 | 1 | 1 | |||
| Всего 14 | 3 | 0 | 5 | 0 | 6 | 6 |
ГЛИЦИН
4.7 Графическое изображение количества глицина в плазме крови спортсменов разных видов спорта и в зависимости от уровня квалификации:


Глицин (мкМ):

Глицин (%).
Таблица 4. 7 Корреляционные связи у борцов, ЛА и хоккеистов (БОРЦЫ, ЛЕГКОАТЛЕТЫ, ХОККЕИСТЫ):
| Показатели | Гли(относит. К-во) | Гли(абсолют. К-во) | Гли(относит к-во) | Гли(абсолют. К-во) | Гли(относит к-во) | Гли(абсолют. К-во) |
| возраст | -,829** | -,705* | ,841** | |||
| рост | ,852* | |||||
| вес | ,913* | |||||
| ип | -,855* | |||||
| длтул | ,900* | |||||
| ширтаза | -,668* | |||||
| длруки | ,845* | |||||
| длног | ,874* | |||||
| отнжира | -,919* | |||||
| отнкостей | ,744* | ,738* | -,829* | |||
| весорг | ,995** | |||||
| кум | ,999* | |||||
| саннастр | -,779* | |||||
| сэу | 1,000** | 1,000** | ,954* | |||
| SpO2 | ,863* | |||||
| NISP | -,718* | |||||
| NIDP | -,767** | |||||
| @__S_ | -,814* | |||||
| TrxA | ,848* | |||||
| ** Корреляция значима на уровне 0.01 (2-сторон.) | ||||||
| * Корреляция значима на уровне 0.05 (2-сторон.). | ||||||
ОБЩЕЕ КОЛИЧЕСТВО:
| глицин | борцы | ЛА | хоккей | |||
| коррел | В т.ч. отр | коррел | В т.ч. отр | коррел | В т.ч.отр | |
| Морфметрия | 5 | 3 | 8 | 2 | 2 | 1 |
| Кардиореспираторная система | 2 | 2 | 4 | 2 | ||
| Психологические особенности | 1 | 1 | 2 | 1 | ||
| Всего 25 | 8 | 6 | 10 | 2 | 7 | 3 |
У всех его много, наиболее стабилен у ЛА (Графики 4.7). Во всех видах спорта встречается, влияя как на сосуды, так и на утилизацию кислорода (Табл 4.7). Не можем признать глицин стержнем, формирующим вид спорта. Но очевидно оказывает неспецифические благотворные воздействия на всех спортсменов.
АЛАНИН
4.8 Графическое изображение количества аланина в плазме крови спортсменов разных видов спорта и в зависимости от уровня квалификации:

Аланин (мкМ):
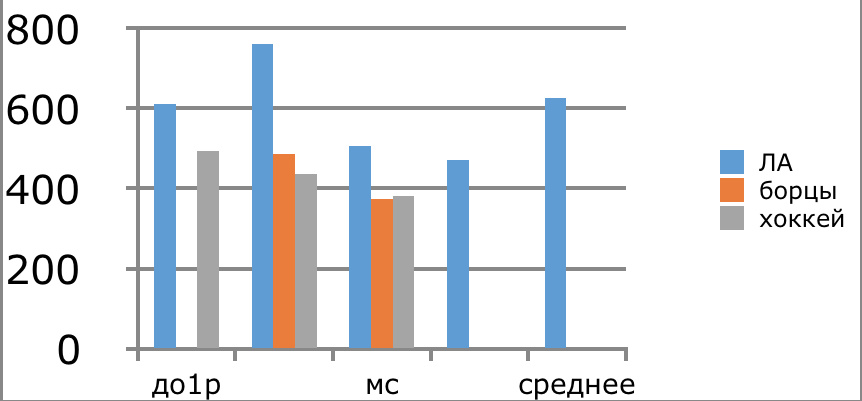
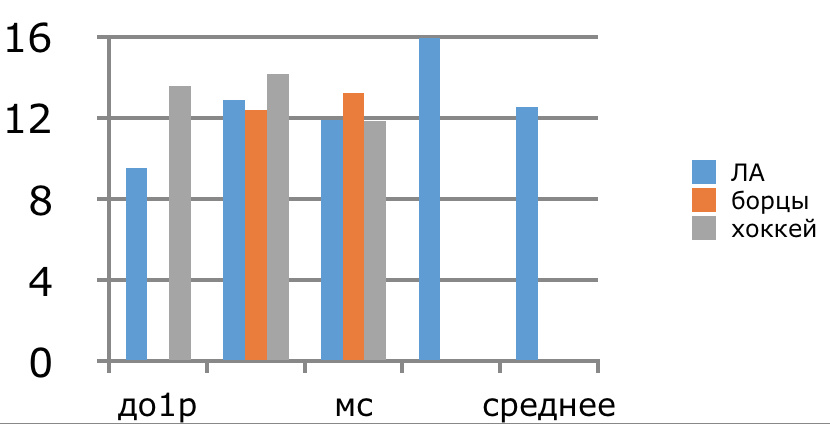
Аланин (%).
Аланин входит в систему детоксикации молочной кислоты. У борцов все корреляции содержания аланина в крови отрицательны. Аланин снижает и межтеменной размер, удельную мощность, эмоциональную устойчивость. Не нужен аланин борцам. Не является частью механизмов, адаптирующих к борьбе.
У легкоатлетов содержание аланина коррелирует положительно с длиннотными размерами – рост, длина туловища, длина конечностей, а также с костным компонентом (Графики 4.8). Аланин влияет на костномозговое кроветворение, то есть улучшает аэробность(Табл.4.8). Нужен легкоатлетам на стадии формирования структуры тела. Помимо глутамина стрежневой аминокислотой для легкоатлетов является аланин. Как маркер детоксикации молочной кислоты, ее метаболизирования. Есть смысл давать на стадии 1 разрядников и КМС. Это фактор борьбы с собственным телом. Борцам и хоккеистам смысла давать нет.
Таблица 4. 8 Корреляционные связи у борцов, ЛА и хоккеистов (БОРЦЫ, ЛЕГКОАТЛЕТЫ, ХОККЕИСТЫ):
| Показатели | Ала(относит. К-во) | Ала(абсолют к-во) | Ала(относит к-во) | Ала(абсолют. К-во) | Ала(относит. К-во) | Ала(абсолют.к-во) |
| возраст | ,928** | |||||
| рост | ,978** | |||||
| ип | -,685* | |||||
| длтул | ,964** | |||||
| длруки | ,866* | |||||
| длног | ,950** | |||||
| весжира | -,676* | |||||
| Вескостей | ,941** | |||||
| Отнкостей | ,894* | |||||
| весорг | -,759* | |||||
| межтем | -,721* | |||||
| пвцудель | -,734* | |||||
| экстраверт | ,959* | |||||
| сэу | -,784* | -1,000** | -1,000** | |||
| ПМР | -1,000** | -1,000** | ,985* | |||
| РСО | -1,000** | -1,000** | ,985* | |||
| NISP | -,863** | 1,000** | -1,000** | |||
| ** Корреляция значима на уровне 0.01 (2-сторн.) | ||||||
| * Корреляция значима на уровне 0.05 (2-сторон.). | ||||||
Аланин ригидность у легкоатлетов снижает, а у хоккеистов – повышает.
ОБЩЕЕ КОЛИЧЕСТВО:
| аланин | борцы | ЛА | хоккей | |||
| коррел | В т.ч. отр | коррел | В т.ч. отр | коррел | В т.ч.отр | |
| Морфметрия | 3 | 3 | 7 | 1 | 1 | |
| КАРДИОРЕСПИРАТОРНАЯ СИСТЕМА | 2 | 2 | ||||
| Психологические особенности | 1 | 1 | 2 | 2 | ||
| Всего 18 | 6 | 6 | 9 | 0 | 3 | 1 |
ВАЛИН
Графическое изображение количества валина в плазме крови спортсменов разных видов спорта и в зависимости от уровня квалификации:
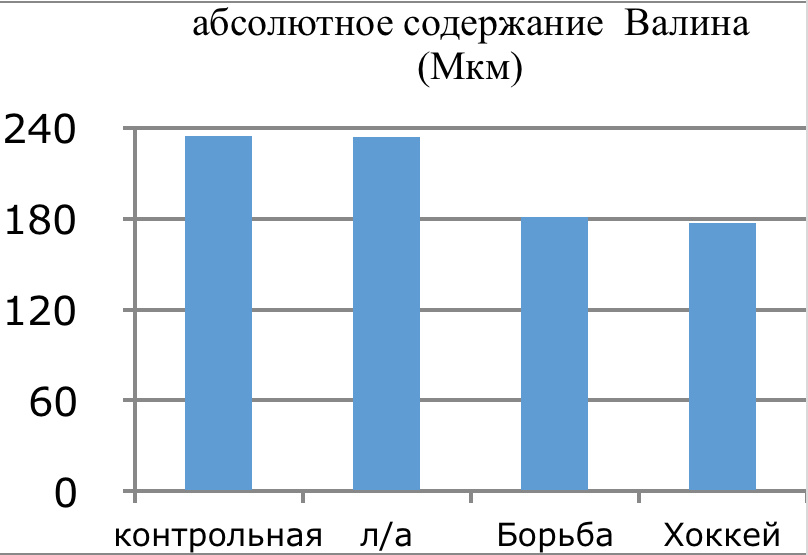
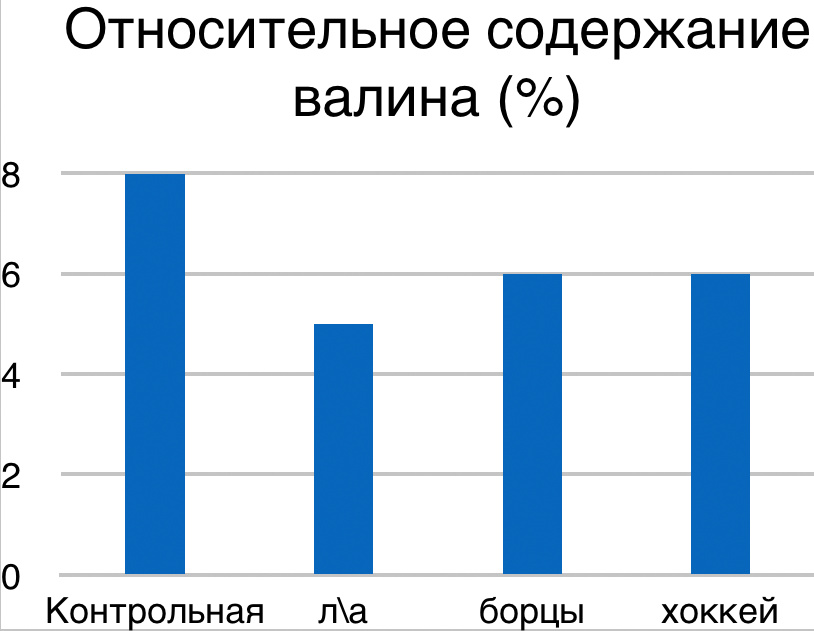
Валин (Мкм):
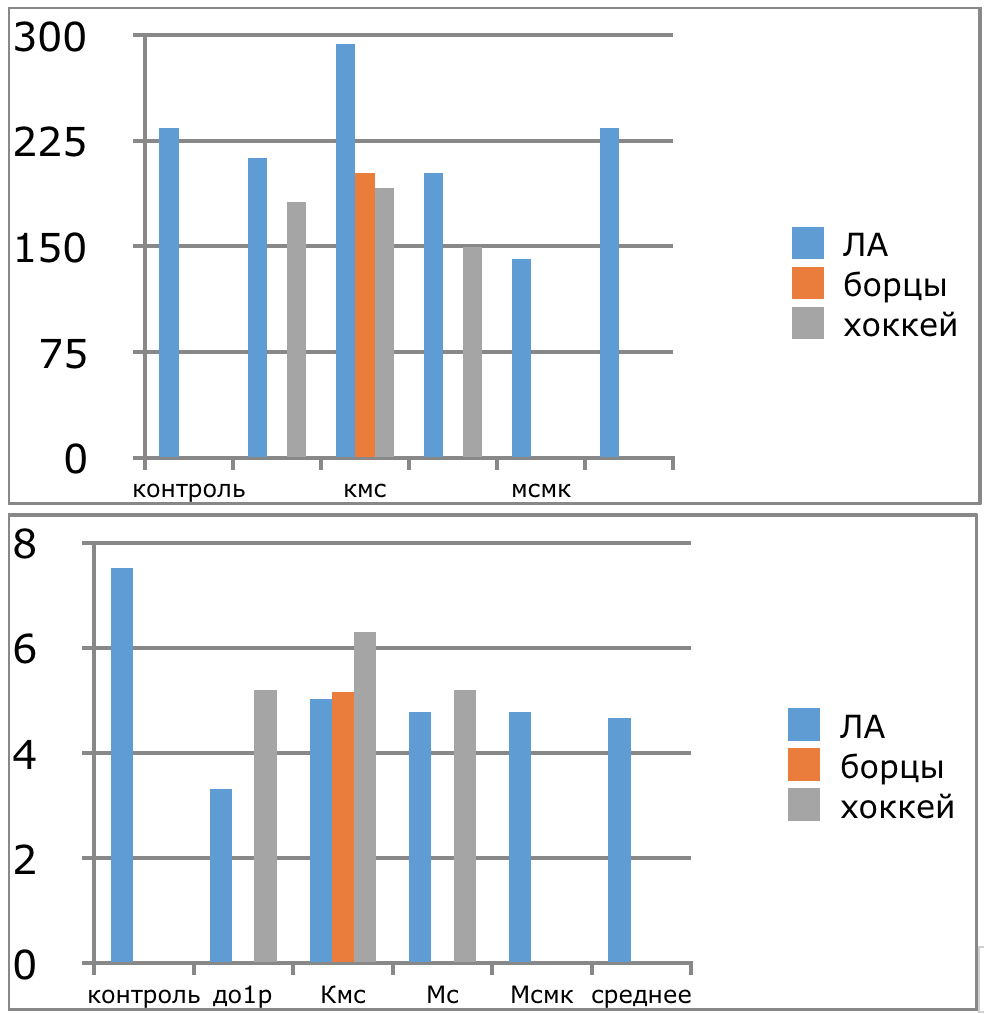
Валин (%).
У борцов абсолютное количество валина меньше, чем в контроле, но относительное содержание валина у борцов от контроля достоверно не отличается( Графики 4.9). С ростом мастерства абсолютное количество валина у борцов снижается, но относительное – возрастает. У борцов содержание валина связано с психологическими и физиологическими параметрами. Повышая ЦВД относительное содержание валина снижает минутный объем, что соответствует закону Франка-Старлинга. Валин снижает экстраверсию, то есть снижает преобладание правого полушария. При этом усиливает РСО, то есть склонность к фиксированным формам поведения в экстремальных ситуациях, что тоже вполне естественно (Табл.4.9).
У легкоатлетов абсолютное содержание валина неотличимо от контроля, но относительное содержание валина – в полтора раза ниже. Тем не менее абсолютное количество валина делает туловище легкоатлета короче и шире, а относительное – делает его менее ригидным и более смелым, но эмоционально неустойчивым. С ростом мастерства вдвое снижается абсолютное количество валина и имеется тенденция к снижению относительного содержания валина.
У хоккеистов и абсолютное и относительное содержание валина ниже, чем в контроле. А с ростом мастерства содержание валина, как и у легкоатлетов, возрастает до уровня КМС, затем снижается. Влияет валин на нейротизм (отрицательно) то есть повышает контроль левого полушария, повышает настроение и соревновательную эмоциональную устойчивость, и тонус крупных сосудов.
Таким образом, содержание валина у хоккеистов («печеночников») влияет больше на взаимоотношение отделов мозга (в т.ч. полушарий) и гемодинамику, а у легкоатлетов( «мозговиков»), кроме того принимает участие в формировании структурного следа адаптации.
Таблица 4. 9 Корреляционные связи у борцов, ЛА и хоккеистов (БОРЦЫ, ЛЕГКОАТЛЕТЫ, ХОККЕИСТЫ):
| Показатели | Вал(относит к-во) | Вал(абсолют. К-во) | Вал(относит. К-во) | Вал(абсолют.к-во) | Вал(относит. К-во) | Вал(абсолют. К-во) |
| Отндлтул | -,962** | |||||
| Ширтаза | ,935** | |||||
| Экстраверт | -,844** | |||||
| Нейротизм | -,988* | |||||
| Саннастр | ,999* | |||||
| Сэу | -1,000** | ,971* | ||||
| СКР | -,978* | |||||
| РСО | ,840* | -1,000** | ||||
| TrxA | -,839* | |||||
| Fw_ ЦВД | ,891** | |||||
| CO_ | -,640* | |||||
| ** Корреляция значима на уровне 0.01 (2-сторн.) | ||||||
| * Корреляция значима на уровне 0.05 (2-сторон.). | ||||||
ОБЩЕЕ КОЛИЧЕСТВО:
| Валин | Борцы | ЛА | хоккей | |||
| коррел | В т.ч. отр | Коррел | В т.ч. отр | коррел | В т.ч.отр | |
| Морфометрия | 2 | 1 | ||||
| Кардиореспираторная система | 2 | 1 | 1 | 1 | ||
| Психологические особенности | 2 | 1 | 1 | 1 | 3 | 1 |
| Всего 11 | 4 | 2 | 3 | 2 | 4 | 2 |
ЦИСТЕИН
4.10 Графическое изображение количества цистеина в плазме крови спортсменов разных видов спорта и в зависимости от уровня квалификации:
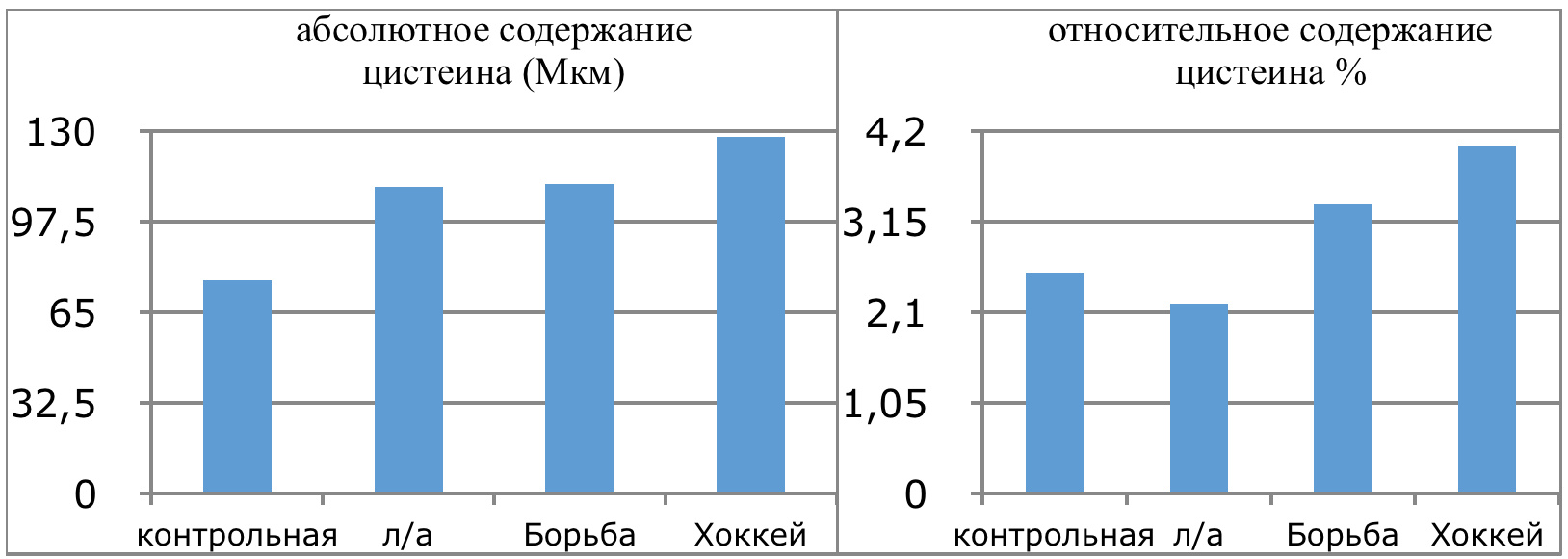
Цистеин (Мкм):
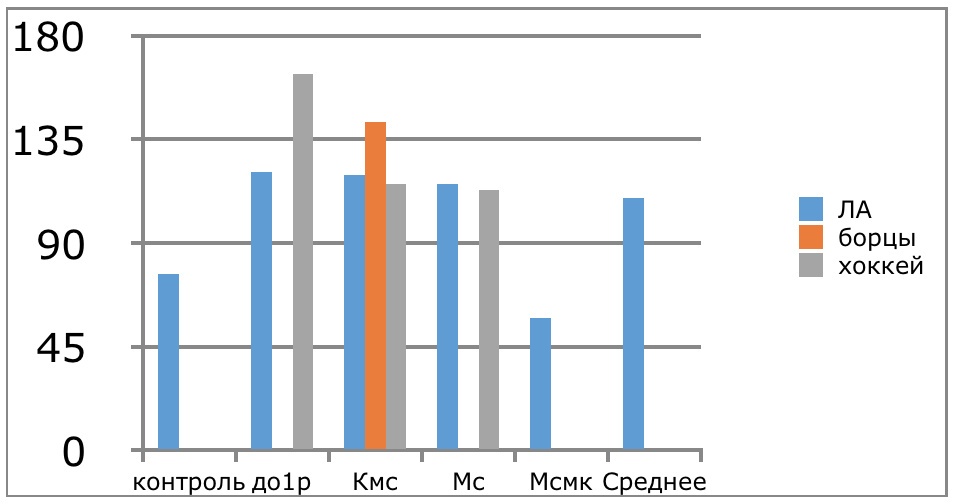
Цистеин (%).
Абсолютное содержание цистеина у представителей рассматриваемых видов спорта выше, чем в контроле. Относительное содержание выше, чем в контроле у «печеночников» – борцов и хоккеистов (Графики 4.10).
У них содержание цистеина снижается с ростом квалификации, не снижаясь при этом до уровня контроля. У легкоатлетов «мозговиков» количество цистеина на уровне МСМК снижается ниже контроля.
У борцов содержание цистеина отрицательно связано с размерами тела, в том числе с длиной ног (Табл.4.10). Кроме того, способствует нейротизму, то есть снижает контроль левого полушария над лимбической системой и снижает эмоциональную устойчивость на соревнованиях. Предположительно за счет того, что у борцов ослаблен переход цистеина в таурин. Кроме того, цистеин снижает артериальное давление, уменьшает фракцию выброса при повышении индекса доставки кислорода. То есть цистеин у борцов тормозит формирование структурного следа адаптации, являясь антагонистом аспарагиновой кислоты.
У легкоатлетов цистеин не влияет на формирование тела (структурного следа адаптации). Содержание цистеина у легкоатлетов положительно коррелирует с экстраверсией, то есть способствует преобладанию правого полушария – уровень локомоций по Бернштейну. Снижая таким образом роль левого полушария снижает эмоциональную соревновательную устойчивость, и ригидность, как свойство личности. Снижает напряжение содержания кислорода, кровяное давление и индекс пациента.
У хоккеистов цистеин способствует тонусу крупных сосудов и ригидности, как свойству личности.
Таким образом, цистеин только у борцов связан с морфологическими параметрами, и у всех – с психологическими.
Максимальное кол-во корреляций с морфологией.
Таблица 4. 10 Корреляционные связи у борцов, ЛА и хоккеистов (БОРЦЫ, ЛЕГКОАТЛЕТЫ, ХОККЕИСТЫ):
| Показатели | Цис(относит.к-во) | Цис(абсолют. К-во) | Цис(относит. К-во) | Цис(абсолют. К-во) | Цис(относит к-во) | Цис(абсолют к-во) |
| Рост | -,723* | |||||
| Ширтаза | -,790* | |||||
| Отнширтаза | -,731* | |||||
| Длног | -,669* | |||||
| Окрголов | -,834** | |||||
| Межтем | -,677* | |||||
| Экстраверт | ,936* | |||||
| Нейротизм | ,757* | |||||
| Сэу | -,785* | -1,000** | ||||
| АР | -1,000** | ,995** | ||||
| PI_ | -,939* | |||||
| SpO2 | -,919* | |||||
| NISP | -,796** | -1,000** | ||||
| NIDP | -,787** | -1,000** | ||||
| TrxA | ,824* | |||||
| EF_ | -,659* | |||||
| DO2 | ,662* | |||||
| ** Корреляция значима на уровне 0.01 (2-сторон.). | ||||||
| * Корреляция значима на уровне 0.05 (2-сторон.). | ||||||
ОБЩЕЕ КОЛИЧЕСТВО:
| цистеин | Борцы | ЛА | хоккей | |||
| Коррел | В т.ч. отр | коррел | В т.ч. отр | коррел | В т.ч.отр. | |
| Морфометрия | 7 | 6 | ||||
| Кардиореспираторная система | 4 | 3 | 1 | |||
| Психологические особенности | 2 | 1 | 2 | 1 | 1 | |
| Всего 17 | 13 | 10 | 2 | 1 | 2 | 0 |
МЕТИОНИН
4.11 Графическое изображение количества метионина в плазме крови спортсменов разных видов спорта и в зависимости от уровня квалификации:
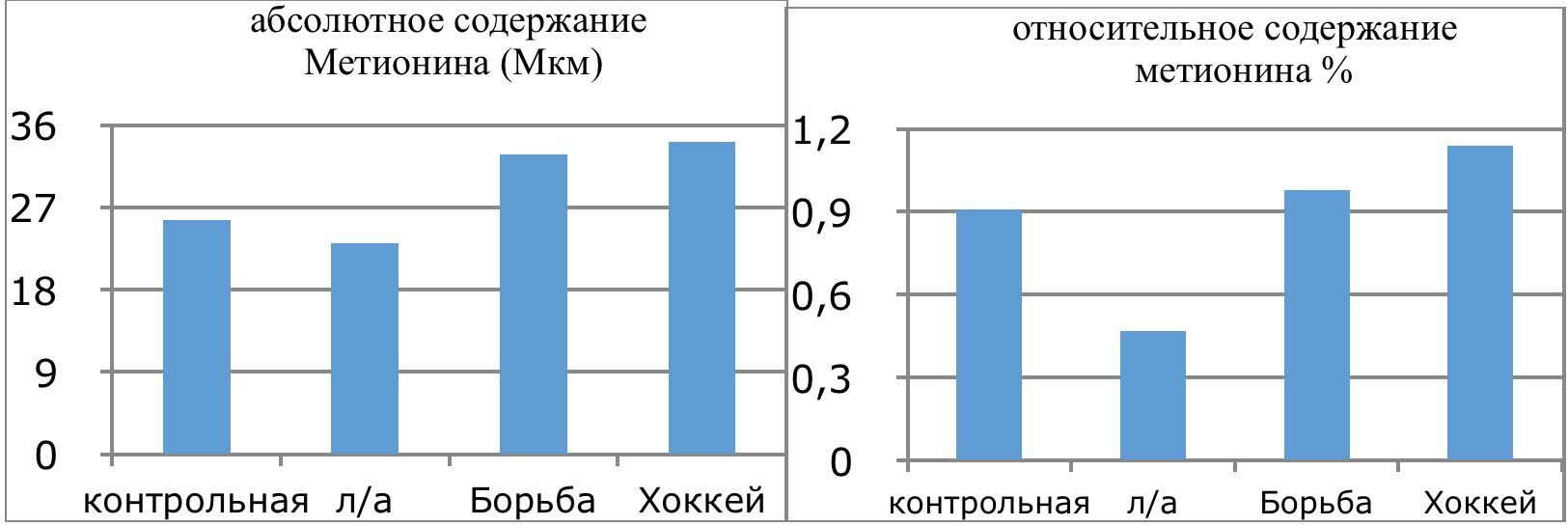
Метионин (Мкм):
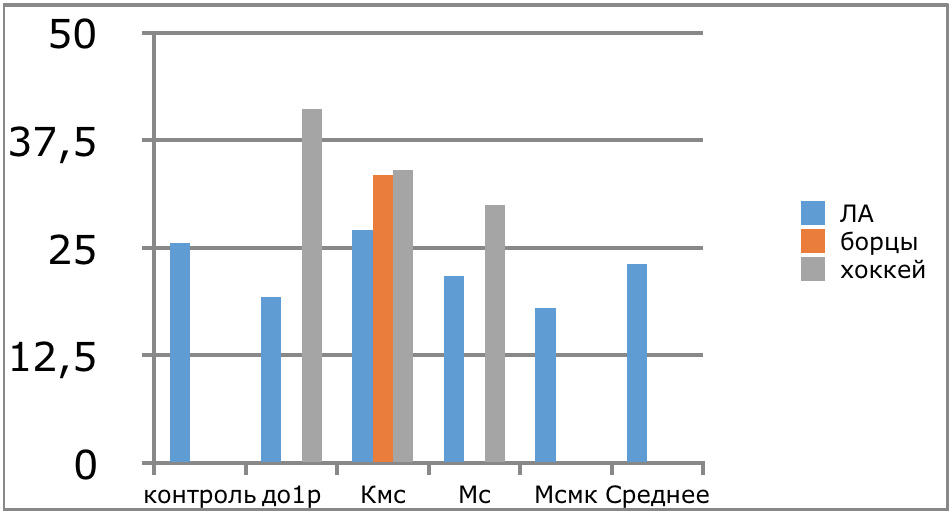
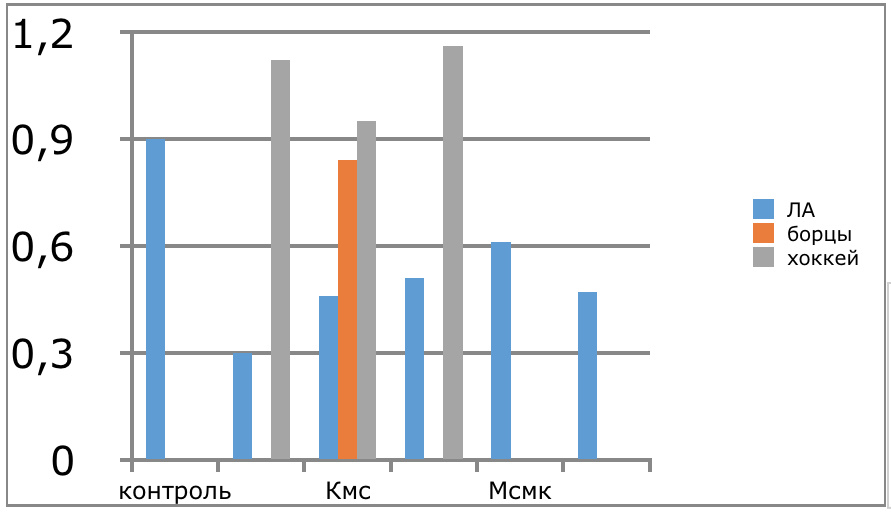
Метионин (%).
Содержание метионина у «печеночников» (борцы и хоккеисты) выше, чем в контроле, а у «мозговиков» (легкоатлеты) – ниже (Графики 4.11).
У борцов относительное количество метионина с ростом квалификации растет. Корреляционные связи только с кислородом крови и тонусом ВНС (положительные) и отрицательная связь с ЧСС. Метионин способствует физиологической боеготовности борца.
У легкоатлетов относительное количество метионина возрастает. Содержание метионина коррелирует с содержанием костного компонента положительно и с длиной рук – отрицательно. Абсолютное количество метионина снижает самочувствие и фракцию выброса и повышает ЧСС(Табл.4.11). Относительное количество метионина повышает активность. Собственно повышение относительного количества метионина способствует активности легкоатлета, отнюдь не повышая работоспособность. Метионин – стартовая готовность легкоатлета, усиливает предстартовую лихорадку. В зависимости от темперамента – холерикам он вреден, меланхоликам – полезен.
Хоккеисты – чемпионы по метионину. У хоккеистов метионин положительно связан с весом тела и связанными с весом показателями – мышечным и костным компонентами, то есть значение метионина по достижении оптимального строения тела снижается, поддерживая только индекс пациента – показатель хорошего самочувствия.
Таблица № 11 Корреляционные связи у борцов, ЛА и хоккеистов (БОРЦЫ, ЛЕГКОАТЛЕТЫ, ХОККЕИСТЫ):
| Показатели | Мет(относит. К-во) | Мет(абсолют. К-во) | Мет(относит. К-во) | Мет(абсолют. К-во) | Мет(относит. К-во) | Мет(абсолют. К-во) |
| Вес | ,762* | |||||
| Весор | ,802* | |||||
| Отндлрук | -,883* | |||||
| Весмышц | ,888** | |||||
| отнмышц | ,764* | |||||
| Вескостей | ,791* | |||||
| Отнкостей | ,903* | |||||
| Межтем | ,932* | |||||
| Сансамоч | -,972* | |||||
| Санактив | ,980* | |||||
| PI_ | ,893* | |||||
| HR_ | -,833** | ,897* | ||||
| SpO2 | ,635* | |||||
| @__S_ | ,715* | |||||
| EF_ | -,882* | |||||
| ** Корреляция значима на уровне 0.01 (2-сторон.). | ||||||
| * Корреляция значима на уровне 0.05 (2-сторон.). | ||||||
ОБЩЕЕ КОЛИЧЕСТВО:
| метионин | борцы | ЛА | хоккей | |||
| коррел | В т.ч. отр | коррел | В т.ч. отр | коррел | В т.ч. отр. | |
| Морфометрия | 3 | 1 | 5 | |||
| Кардиореспираторная система | 3 | 1 | 2 | 1 | ||
| Психологические особенности | 2 | 1 | ||||
| Всего 15 | 3 | 1 | 7 | 3 | 5 | 0 |
ИЗОЛЕЙЦИН
4.12 Графическое изображение количества изолейцина в плазме крови спортсменов разных видов спорта и в зависимости от уровня квалификации:
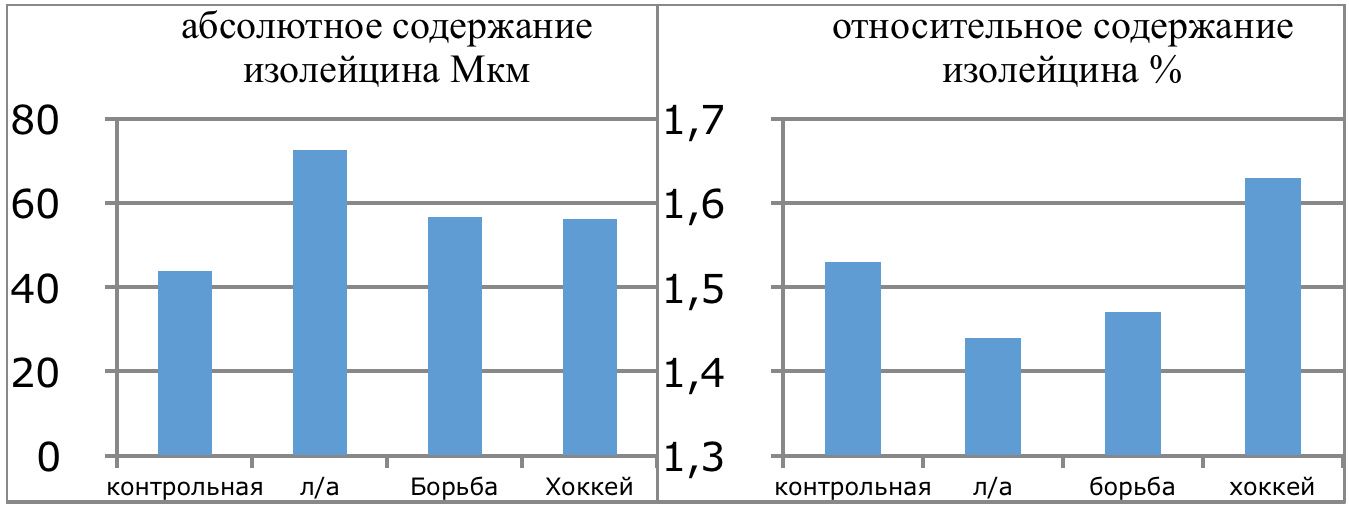
Изолейцин (Мкм):
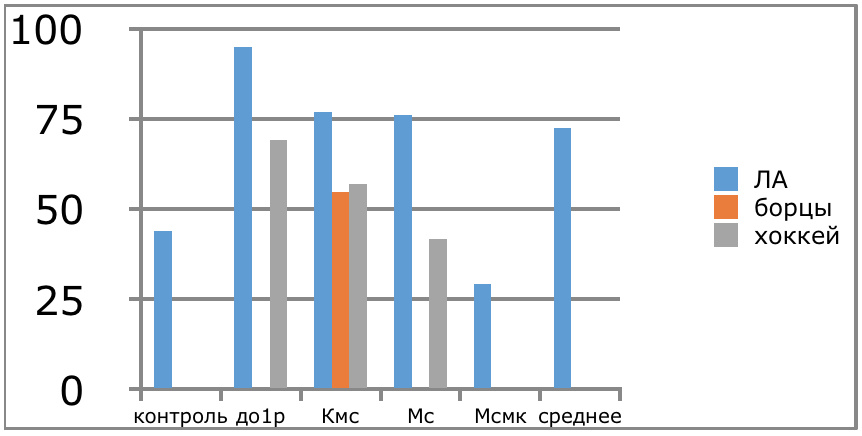
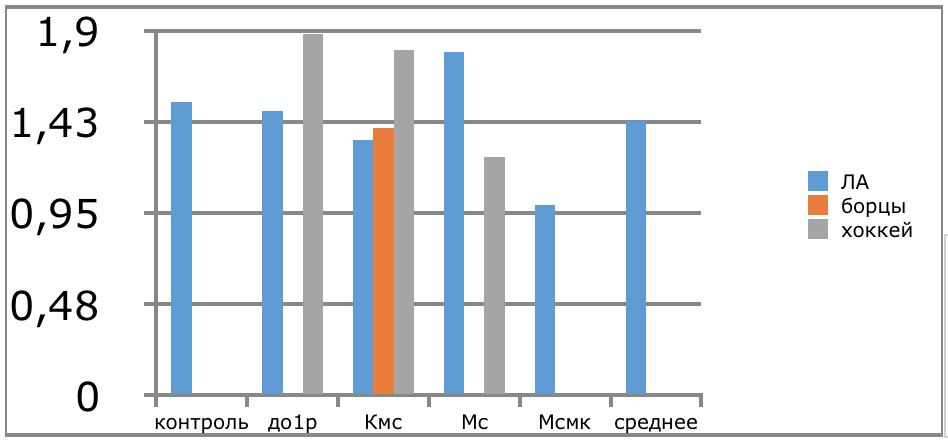
Изолейцин (%).
Уровень абсолютного количества изолейцина в плазме крови у легкоатлетов больше, чем в контроле и у борцов и хоккеистов, а относительное количество – ниже чем в контроле и у борцов и хоккеистов (Графики 4.12).
Для борцов изолейцин фактор снижения возможностей находить решение в экстремальных ситуациях – и только. До уровня мастера спорта изолейцин у борцов возрастает.
У легкоатлетов изолейцин – фактор экстраверсии, то есть усиливает правополушарную активность, следствием чего является повышение мотивации к легкой атлетике (бегу). Косвенно тому же способствует положительная корреляция с относительной длиной ног у легкоатлетов. Резкое падение изолейцина у мастеров спорта международного класса прямо говорит о том, что для легкоатлетов изолейцин является важным фактором, но не для достижения высшего мастерства (Табл. 4.12).
У хоккеистов изолейцин снижается с ростом мастерства, что вполне естественно – эта аминокислота снижает мотивацию заниматься видом спорта, как мы видим – в пользу легкой атлетики. Хотя достоверных корреляций изолейцина с показателями темперамента мы не наблюдаем, но можем предположить, что при расширении исследований мы такие корреляции обнаружим.
Изолейцин – фактор влияющий преимущественно на психологию, и только у легкоатлетов формирует структурный след адаптиции.
Таблица 4.12 Корреляционные связи у борцов, ЛА и хоккеистов (БОРЦЫ, ЛЕГКОАТЛЕТЫ, ХОККЕИСТЫ):
| Показатели | Иле(относит.к-во) | Иле(абсолют. к-во) | Иле(относит. к-во) | Иле(абсолют. К-во) | Иле(относит. К-во) | Иле(абсолют. К-во) | ||
| отндлног | ,824* | |||||||
| экстраверт | ,886* | |||||||
| Мэ | 1,000** | 1,000** | -,977* | -,981* | ||||
| РСО | ,858* | 1,000** | 1,000** | |||||
| Fw_ | ,889* | |||||||
| ** Корреляция значима на уровне 0.01 (2-сторон.). | ||||||||
| * Корреляция значима на уровне 0.05 (2-сторон.). | ||||||||
ОБЩЕЕ КОЛИЧЕСТВО:
| изолейцин | борцы | ЛА | хоккей | |||
| коррел | В т.ч. отр | коррел | В т.ч. отр | коррел | В т.ч.отр | |
| Морфометрия | 1 | |||||
| Кардиореспираторная система | 1 | |||||
| Психологичесмкие особенности | 1 | 1 | 2 | 2 | ||
| Всего 5 | 1 | 0 | 3 | 0 | 2 | 2 |
ЛЕЙЦИН
4.13 Графическое изображение количества лейцина в плазме крови спортсменов разных видов спорта и в зависимости от уровня квалификации:
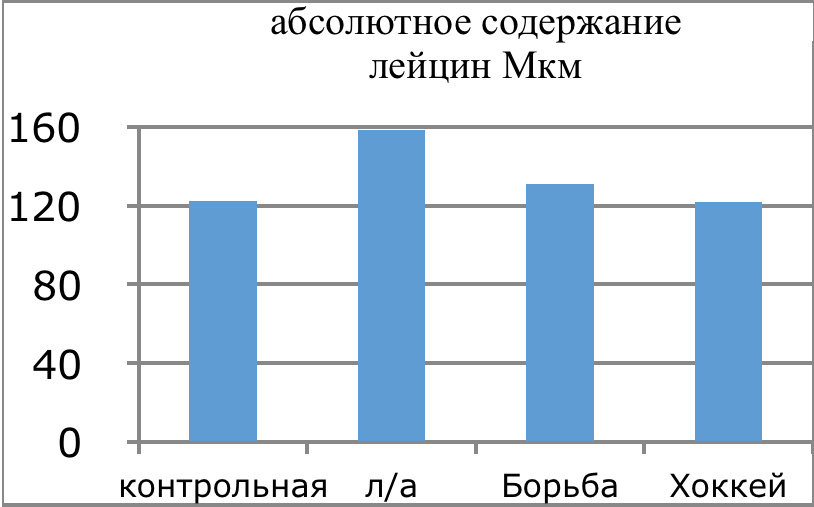
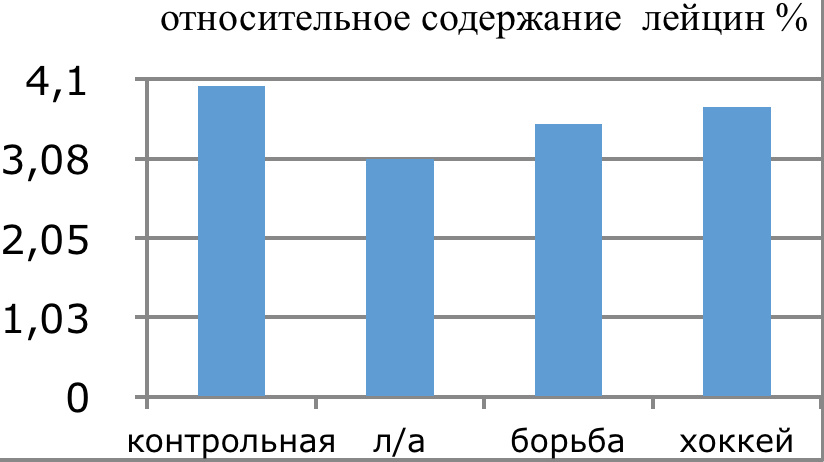
Лейцин (Мкм):
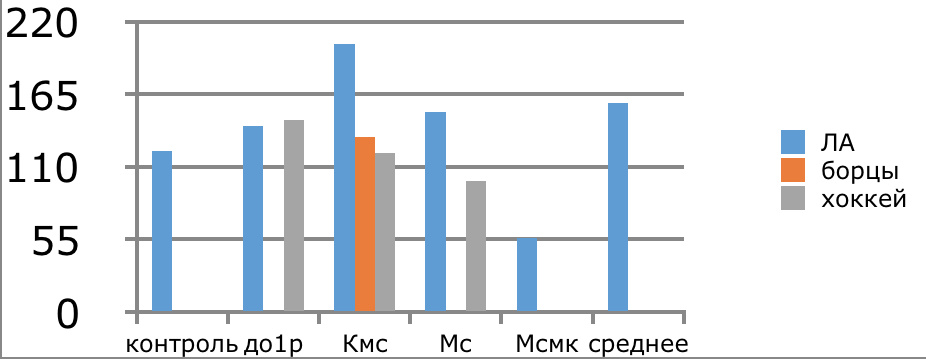
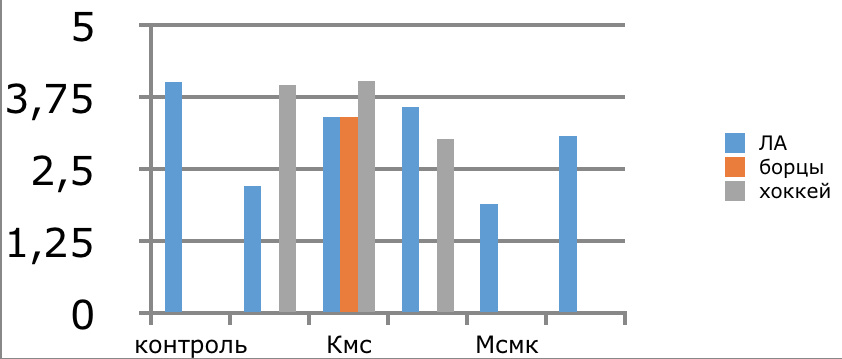
Лейцин (%).
Содержание лейцина в плазме крови спортсменов так же зависит от вида спорта. И снова у легкоатлетов абсолютное количество лейцина больше, чем у других, а относительное – меньше (Графики 4.13).
У борцов среднее содержание лейцина неотличимо от такового в контроле и с ростом квалификации до уровня мастера спорта возрастает выше контроля. Критическая удельная мощность (мощность, при которой происходит срыв дыхания) с ростом уровня лейцина снижается, видимо это и является специфическим для борьбы проявлением лейцина. Снижение индекса пациента и артериального давления можно рассматривать как побочный и вредный эффект. Кроме того, повышение склонности к ФФП до уровня мастера спорта, скорее, не является негативным явлением.
У легкоатлетов содержание лейцина возрастает с ростом мастерства, и резко снижается у мастеров спорта международного класса. Рост содержания лейцина у легкоатлетов укорачивает и расширяет туловище и сопровождается ростом веса жира. Снижает ригидность актуальную и преморбидную. То есть для легкоатлетов в стадии формирования структурного следа адаптации лейцин необходим (Табл.4.13).
У хоккеистов количество лейцина снижается с возрастом, так что снижение с ростом мастерства может быть обусловлено именно этим. Отмечена положительная корреляция с костным компонентом. Сдвиг тонуса ВНС в сторону парасимпатики видимо и является причиной снижения соревновательной эмоциональной устойчивости.
Лейцин связан с психологическими параметрами, с морфологией и физиологией у хоккеистов, а у борцов влияет на психологические и физиологические параметры.
Таблица 4.13 Корреляционные связи у борцов, ЛА и хоккеистов (БОРЦЫ, ЛЕГКОАТЛЕТЫ, ХОККЕИСТЫ):
| Показатели | Лей(относит. К-во) | Лей(абсолют. К-во) | Лей(относит. К-во) | Лей(абсолют. К-во) | Лей(относит. К-во) | Лей(абсолют. К-во) |
| возраст | -,806* | |||||
| отндлтул | -,825* | |||||
| ширтаза | ,831* | |||||
| весжира | ,948* | |||||
| отнкостей | ,806* | |||||
| кум | -,785* | |||||
| сэу | -,977* | |||||
| АР | -,951* | |||||
| ПМР | -,989* | |||||
| РСО | ,956** | |||||
| PI_ | -,739* | |||||
| NISP | -,634* | |||||
| @__S_ | -,838* | |||||
| ** Корреляция значима на уровне 0.01 (2-сторон.). | ||||||
| * Корреляция значима на уровне 0.05 (2-сторон.). | ||||||
ОБЩЕЕ КОЛИЧЕСТВО:
| лейцин | борцы | ЛА | хоккей | |||
| коррел | В т.ч. отр | коррел | В т.ч. отр | коррел | В т.ч. отр | |
| Морфометрия | 3 | 1 | 2 | 1 | ||
| Кардиореспираторная система | 3 | 3 | 1 | 1 | ||
| Психологические особенности | 1 | 2 | 2 | 1 | 1 | |
| Всего 13 | 4 | 3 | 5 | 3 | 4 | 3 |
ТИРОЗИН
4.14 Графическое изображение количества тирозина в плазме крови спортсменов разных видов спорта и в зависимости от уровня квалификации:
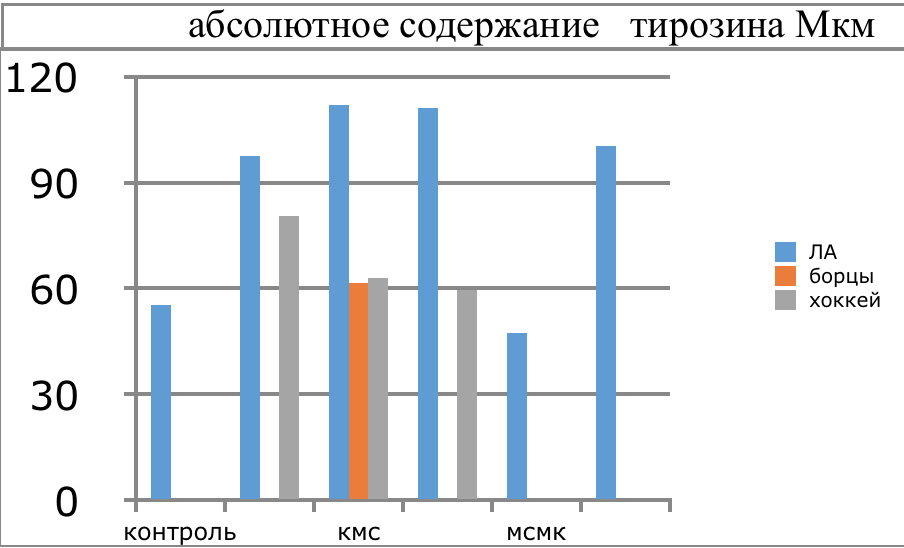
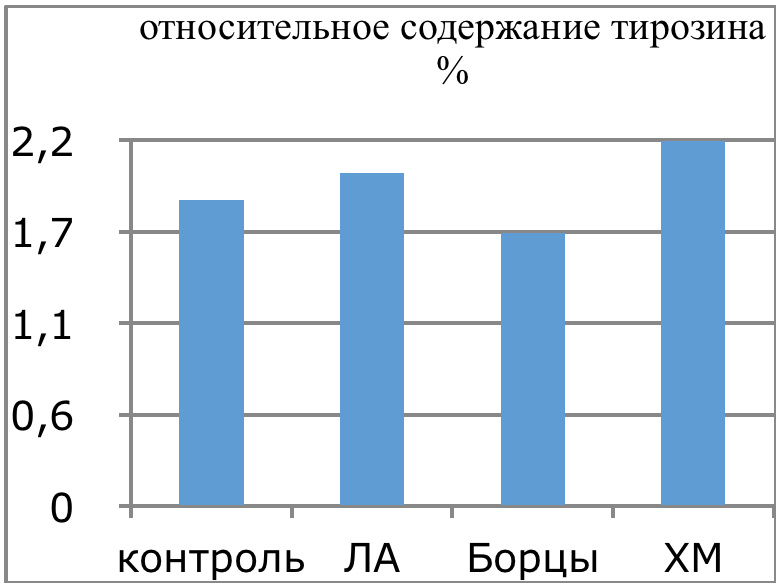
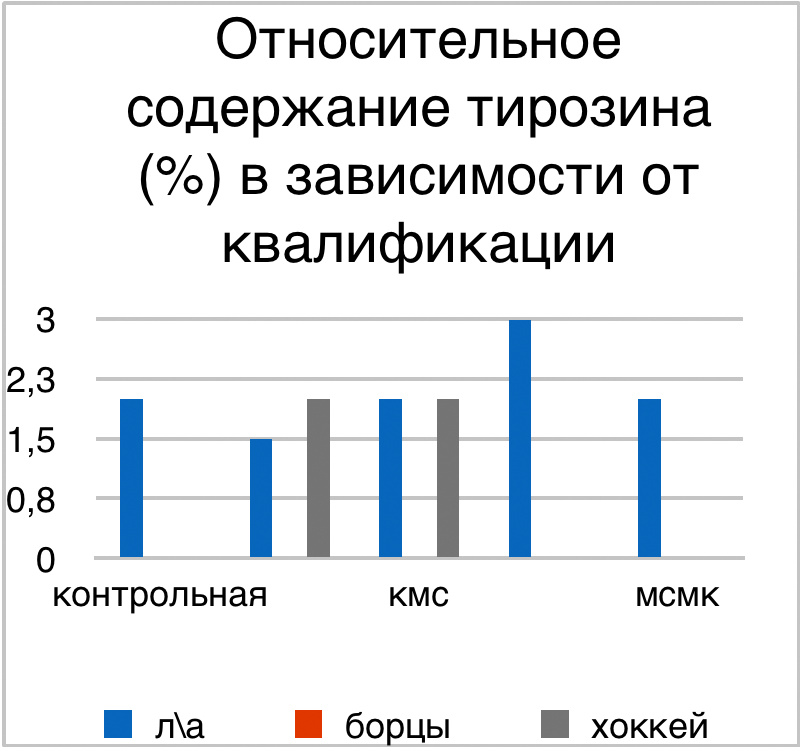
Абсолютное количество тирозина у борцов и хоккеистов не имеет достоверных отличий от контроля. У легкоатлетов абсолютное количество тирозина почти вдвое выше. Относительное количество тирозина у спортсменов одинаково с контролем (Графики 4.14).
Метаболизм тирозина сосредоточен в печени (ТАТ – тирозинаминотрансфераза) и регулируется глюкокортикоидами. Поэтому «печеночники» (борцы и хоккеисты) легко снижают уровень тирозина, а «мозговики» (легкоатлеты) могут это делать только на уровне МСМК.
У борцов на строение тела не влияет, влияет на мотивацию – отрицательно (Табл.4.14).
Таблица 4. 14 Корреляционные связи у борцов, ЛА и хоккеистов:
| БОРЦЫ | ЛЕГКОАТЛЕТЫ | ХОККЕИСТЫ | ||||
| Показатели | Тир(относит. К-во) | Тир(абсолют. К-во) | Тир(относит. К-во) | Тир(абсолют. К-во) | Тир(относит. К-во) | Тир(абсолют. К-во) |
| вес | ,716* | |||||
| весор | ,743* | |||||
| отнширпл | -,831* | |||||
| ширтаза | ,872* | |||||
| отнширтаза | ,896* | |||||
| отнжира | ,966** | |||||
| вескостей | ,761* | |||||
| перзад | ,983** | |||||
| пвцудель | ,998* | |||||
| сэу | -1,000** | -1,000** | ,997** | |||
| мэ | -,783* | -1,000** | -1,000** | |||
| PI_ | -,749* | |||||
| NISP | -,742* | |||||
| CO_ | ,661* | |||||
| CI_ | ,679* | |||||
| DO2 | ,705* | |||||
| ** Корреляция значима на уровне 0.01 (2-сторон.). | ||||||
| * Корреляция значима на уровне 0.05 (2-сторон.). | ||||||
ОБЩЕЕ КОЛИЧЕСТВО:
| тирозин | борцы | ЛА | хоккей | |||
| коррел | В т.ч. отр | коррел | В т.ч. отр | коррел | В т.ч.отр | |
| Морфометрия | 5 | 3 | ||||
| КАРДИОРЕСПИРАТОРНАЯ СИСТЕМА | 5 | 2 | 1 | |||
| Психологические особенности | 1 | 1 | 1 | 1 | ||
| Всего 17 | 6 | 3 | 6 | 0 | 5 | 0 |
ФЕНИЛАЛАНИН
Фенилаланин – аминокислота являющаяся предшественником адреналовых и тиреоидных гормонов поэтому не удивительно, что она имеет наибольшее количество корреляционных связей с физиологическими и психологическими особенностями у всех исследуемых спортсменов (Табл.4.15). Но наибольшее количество фенилаланина мы наблюдаем у легкоатлетов, что видимо связано с особеностями их стрессовой реакции на тренировочные и соревновательные нагрузки.
4.15 Графическое изображение количества фенилаланиана в плазме крови спортсменов разных видов спорта и в зависимости от уровня квалификации:
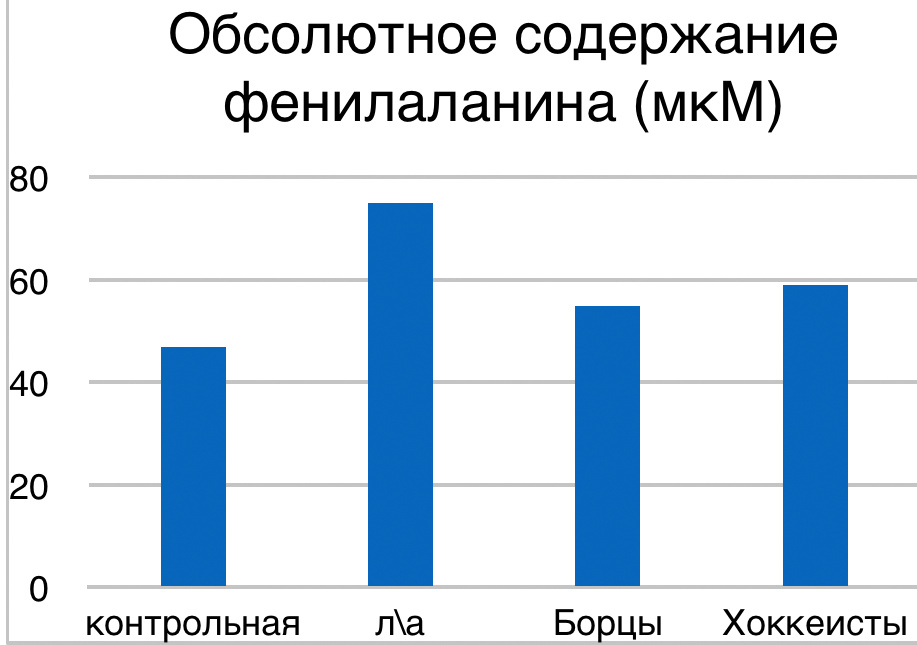
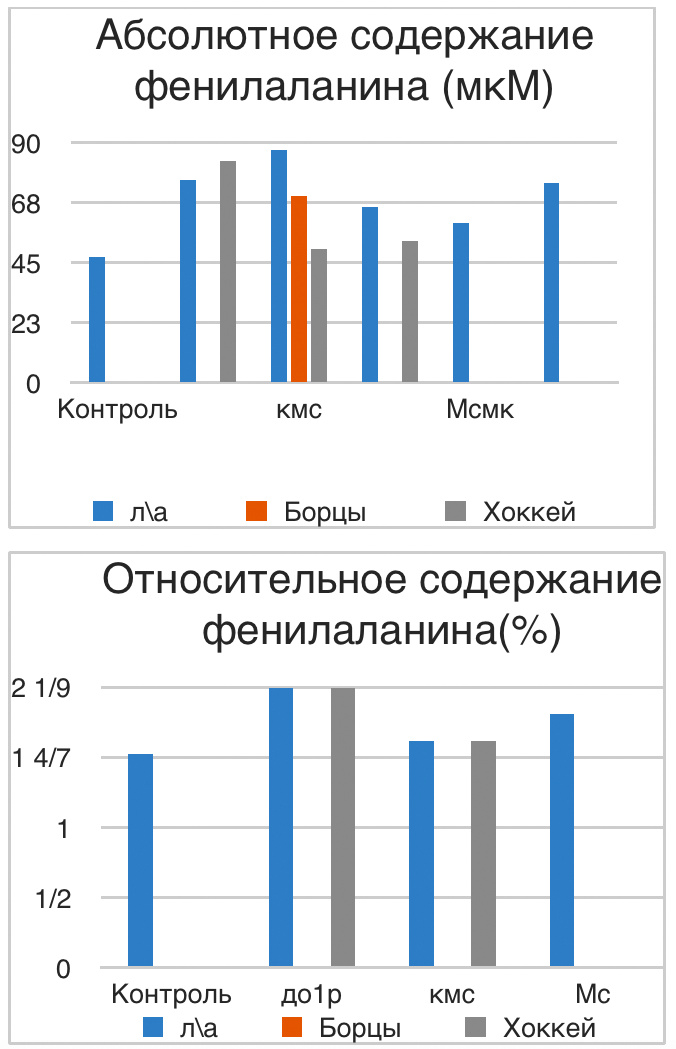
Таблица 4. 15 Корреляционные связи у борцов, ЛА и хоккеистов:
| БОРЦЫ | ЛЕГКОАТЛЕТЫ | ХОККЕИСТЫ | ||||
| Показатели | Фен(относит. К-во) | Фен(абсолют. К-во) | Фен(относит. К-во) | Фен(абсолют. К-во) | Фен(относит. К-во) | Фен(абсолют. К-во) |
| отндлног | -,910* | |||||
| весорг | -,945* | |||||
| нейротизм | -,999** | |||||
| саннастр | ,999* | |||||
| сэу | -1,000** | -1,000** | ,981* | |||
| АР | -,994** | |||||
| РСО | ,831* | -1,000** | -1,000** | |||
| торзсенс | ,918** | 1,000** | 1,000** | |||
| ToeA | -,932* | |||||
| NISP | -,764* | 1,000** | -1,000** | |||
| @__S_ | -,879* | -,814* | ||||
| ** Корреляция значима на уровне 0.01 (2-сторон.). | ||||||
| * Корреляция значима на уровне 0.05 (2-сторон.). | ||||||
ОБЩЕЕ КОЛИЧЕСТВО:
| фенилаланин | борцы | ЛА | хоккей | |||
| коррел | В т.ч. отр | коррел | В т.ч. отр | коррел | В т.ч. отр. | |
| Морфометрия | 2 | 2 | ||||
| Кардиореспираторная система | 1 | 1 | 2 | 2 | 1 | 1 |
| Психологические особенности | 2 | 3 | 3 | 3 | 1 | |
| Всего 12 | 1 | 1 | 7 | 7 | 4 | 2 |
ТРИПТОФАН
Фермент триптофанмонооксигеназа в мозге направляет метаболизм триптофана по серотониновому пути – 5%, второй путь триптофандиоксигеназа – в печени – по кинуренинновому пути – 95%, видимо поэтому у легкоатлетов количество триптофана возрастает до мастеров спорта. А у МСМК резко проваливается, то есть участвует в оксидативных процессах при интенсивном метаболизировании триптофана по кикурениновому пути (Графики 4.16).
У всех спортсменов в той или иной степени имеется хронический стресс[197] (реконструктивный протеолиз). Причем легкоатлеты, в отличии от борцов и хоккеистов, справится с ним долго не могут. Триптофан – маркер легкоатлетов.
У борцов количество триптофана резко снижено во всех исследуемых группах. Скорее всего это связано с тем, что у борцов преобладает «печеночный» адаптивный тип и они интенсивно метаболизируют триптофан по кинурениновому пути. Триптофан через диоксигеназу превращается в витамин РР и нуклеотиды, что не исключает серотонинового пути в мозге, через триптофанмонооксигеназу. Недостаток печени влечет за собой высокий уровень триптофана, и снижает настроение и самочувствие. Поэтому у борцов снижение количества триптофана даже не в 4 раза как у легкоатлетов, а на порядок.
У хоккеистов то же отмечается снижение количества триптофана в крови, но более плавное, чем у борцов. То есть у хоккеистов так интенсивно как у борцов мозг не подключается к обмену триптофана (Табл.4.16).
4.16 Графическое изображение количества триптофана в плазме крови спортсменов разных видов спорта и в зависимости от уровня квалификации:
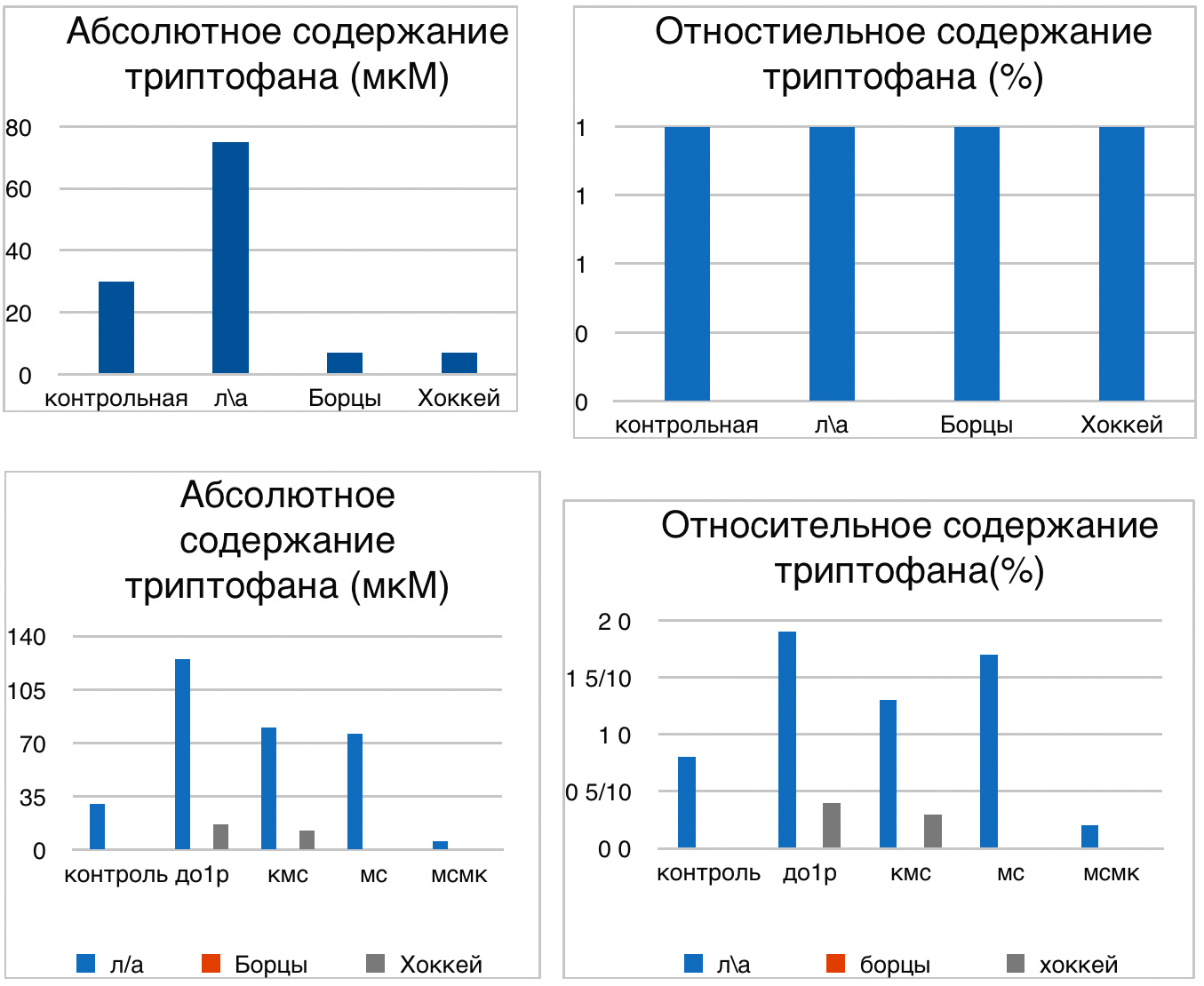
Таблица № 16 Корреляционные связи у борцов, ЛА и хоккеистов:
| Показатели | Три(относит. К-во) | Три(абсолют. К-во) | Три(относит. К-во) | Три(абсолют.к-во) | Три(относит. К-во) | Три(абсолют. К-во) |
| возраст | -,813* | |||||
| отндлрук | -,770* | -,718* | ||||
| сансамоч | -,803* | -,763* | ||||
| санактив | ,987* | ,951* | ||||
| саннастр | -,767* | |||||
| PI_ | -,956* | -,891* | ||||
| HR_ | -,815* | |||||
| SpO2 | -,640* | -,945* | ||||
| SV_ | -,909* | |||||
| DO2 | -,906* | |||||
| ** Корреляция значима на уровне 0.01 (2-сторон.). | ||||||
| * Корреляция значима на уровне 0.05 (2-сторон.). | ||||||
ОБЩЕЕ КОЛИЧЕСТВО:
| триптофан | борцы | ЛА | хоккей | |||
| коррел | В т.ч. отр | Коррел | В т.ч. отр | коррел | В т.ч.отр | |
| Морфометрия | 2 | 2 | 1 | 1 | ||
| Кардиореспираторная система | 1 | 1 | 5 | 5 | 1 | 1 |
| Психологические особенности | 3 | 3 | 2 | 2 | ||
| Всего 15 | 6 | 6 | 8 | 8 | 1 | 1 |
ОРНИТИН
Показатель сохранности мембран печени. Метаболит синтеза мочевины и синтеза полиаминов – азотистые метаболические системы, связанные с детоксикацией и стимуляцией анаболических процессов. Поэтому много корреляций у «печеночников», при этом как следует из полученных результатов — влияет на психику (Табл.4.17).
Борцам он не нужен, должен интенсивно метаболизироваться.
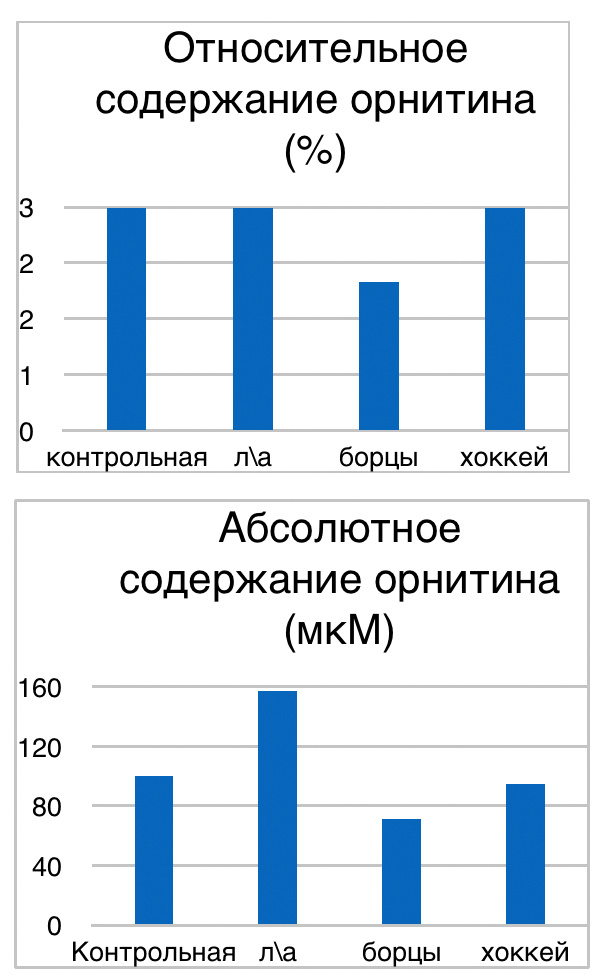
4.17 Графическое изображение количества орнитина в плазме крови спортсменов разных видов спорта и в зависимости от уровня квалификации:
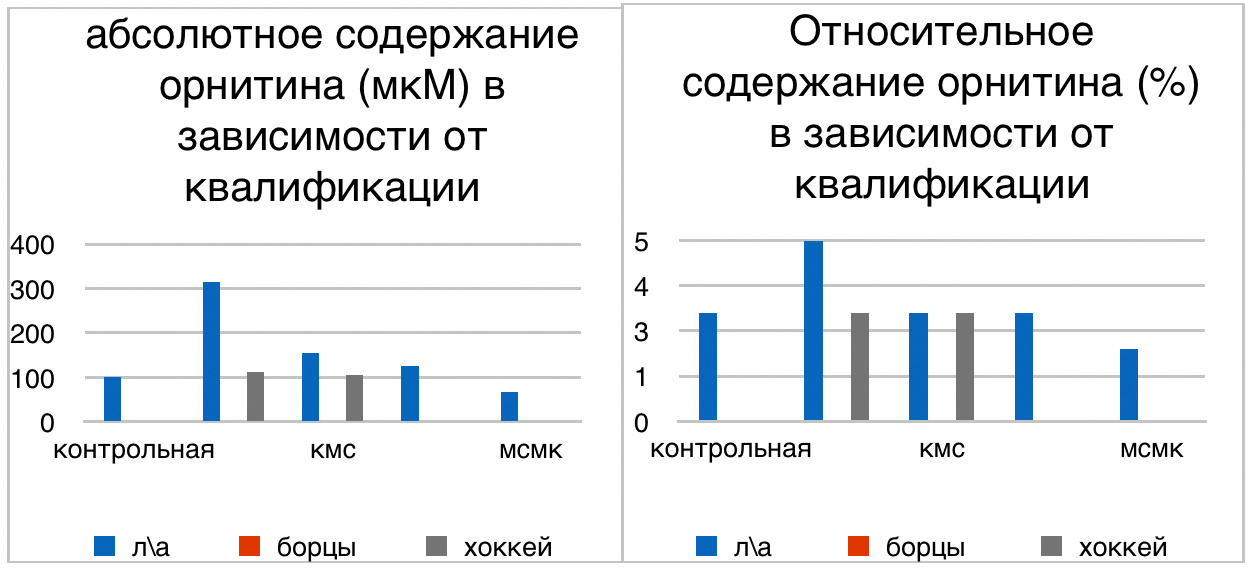
Таблица 4. 17 Корреляционные связи у борцов, ЛА и хоккеистов (БОРЦЫ, ЛЕГКОАТЛЕТЫ, ХОККЕИСТЫ):
| Показатели | ||||||
| Орн(относит. К-во) | Орн(абсолют. К-во) | Орн(относит. К-во) | Орн(абсолют. К-во) | Орн(относит. К-во) | Орн(абсолют. К-во) | |
| ип | -,744* | |||||
| длтул | ,803* | |||||
| отндлтул | ,878** | ,758* | ||||
| ширпл | -,750* | |||||
| длруки | -,767* | |||||
| отндлрук | -,776* | |||||
| окрголов | ,802* | |||||
| перзад | ,801* | |||||
| нейротизм | ,822* | |||||
| мэ | -,857** | 1,000** | 1,000** | |||
| стп | 1,000** | 1,000** | -,996** | |||
| PI_ | -,635* | |||||
| Fw_ | ,761* | |||||
| CO_ | -,740* | |||||
| CI_ | -,706* | |||||
| DO2 | -,669* | |||||
| ** Корреляция значима на уровне 0.01 (2-сторон.). | ||||||
| * Корреляция значима на уровне 0.05 (2-сторон.). | ||||||
ОБЩЕЕ КОЛИЧЕСТВО:
| орнитин | борцы | ЛА | хоккей | |||
| коррел | В т.ч. отр | коррел | В т.ч. отр | коррел | В т.ч. отр | |
| Морфометрия | 4 | 4 | 2 | 5 | ||
| КАРДИОРЕСПИРАТОРНАЯ СИСТЕМА | 5 | 4 | ||||
| Психологические особенности | 2 | 1 | 1 | 1 | 1 | |
| Всего 20 | 11 | 9 | 3 | 0 | 6 | 1 |
ГИСТИДИН
4.18 Графическое изображение количества гистидина в плазме крови спортсменов разных видов спорта и в зависимости от уровня квалификации:
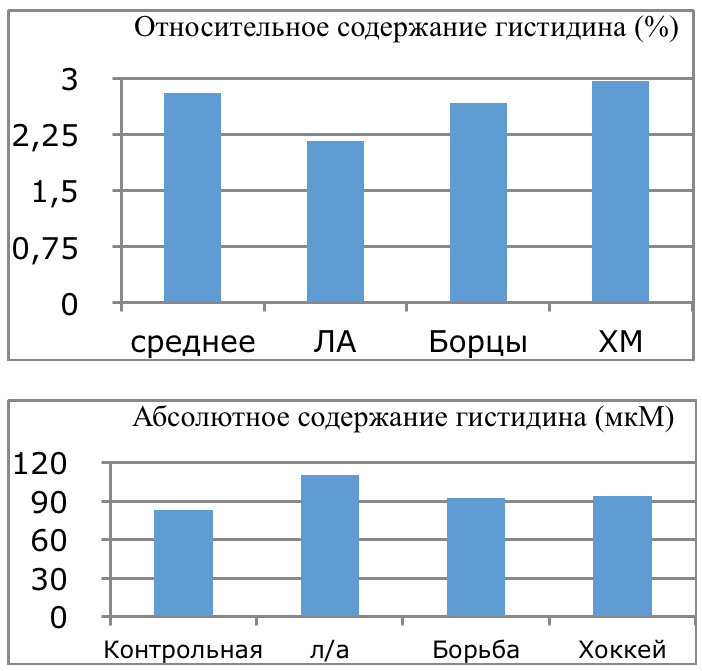
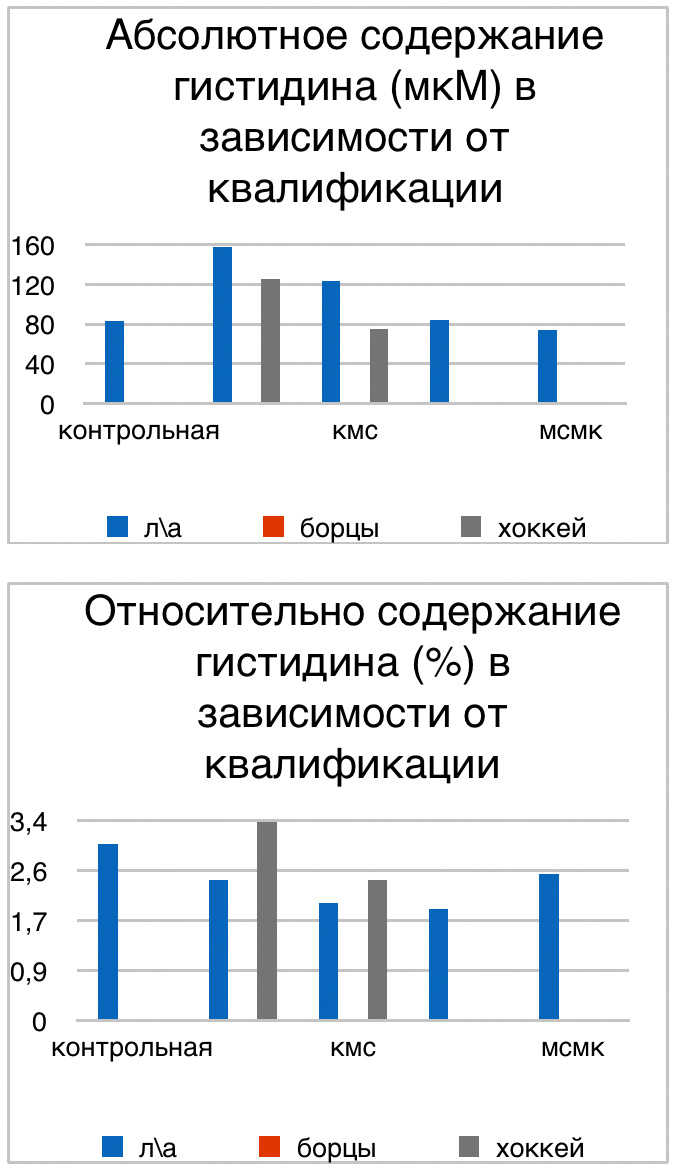
Таблица 4. 18 Корреляционные связи у борцов, ЛА и хоккеистов (БОРЦЫ, ЛЕГКОАТЛЕТЫ, ХОККЕИСТЫ):
| Показатели | Гис(относит. К-во) | Гис(абсолют. К-во) | Гис(относит. К-во) | Гис(абсолют. К-во) | Гис(относит к-во) | Гис)абсолют. К-во) |
| ип | ,947** | |||||
| отндлтул | -,707* | |||||
| ширтаза | -,710* | |||||
| отнширтаза | -,712* | |||||
| длруки | ,821* | |||||
| отндлрук | ,891* | |||||
| весжира | ,769* | |||||
| отнкостей | -,895* | |||||
| отнорг | ,881* | -,777* | ||||
| окрголов | -,684* | -,679* | ||||
| межтем | -,773* | -,966** | ||||
| сансамоч | ,987* | |||||
| санактив | ,959* | |||||
| саннастр | -,999* | |||||
| сэу | -,811* | -1,000** | -1,000** | |||
| мэ | -,749* | -1,000** | -1,000** | |||
| торзсенс | 1,000** | 1,000** | ,957* | |||
| ToeA | ,906* | |||||
| NISP | -,882** | 1,000** | -1,000** | |||
| EF_ | ,884* | |||||
| ** Корреляция значима на уровне 0.01 (2-сторон.). | ||||||
| * Корреляция значима на уровне 0.05 (2-сторон.). | ||||||
ОБЩЕЕ КОЛИЧЕСТВО:
| гистидин | борцы | ЛА | хоккей | |||
| коррел | В т.ч. отр | коррел | В т.ч. отр | коррел | В т.ч.отр | |
| Морфометрия | 5 | 5 | 5 | 2 | 4 | 2 |
| Кардиореспираторная система | 1 | 1 | 2 | 1 | ||
| Психологические особенности | 2 | 2 | 3 | 2 | ||
| Всего 20 | 8 | 8 | 5 | 7 | 7 | 2 |
Гистидин — предшественник гистамина, то есть мощного биологически активного метаболита, выполняющего медиаторные функции. Отсюда вполне закономерные корреляционные связи с физиологическими и психологическими показателями(Табл.4.18). Но согласно полученным нами данным это первейшее средство формирования тела. Ибо максимальное количество корреляционных связей отмечается с морфометрическими показателями. Он не анаболик, он регулятор формы тела, пропорциональности. Его не должно быть много. Это регулятор. Резец скульптора.
АРГИНИН
4.19 Графическое изображение количества аргинина в плазме крови спортсменов разных видов спорта и в зависимости от уровня квалификации:
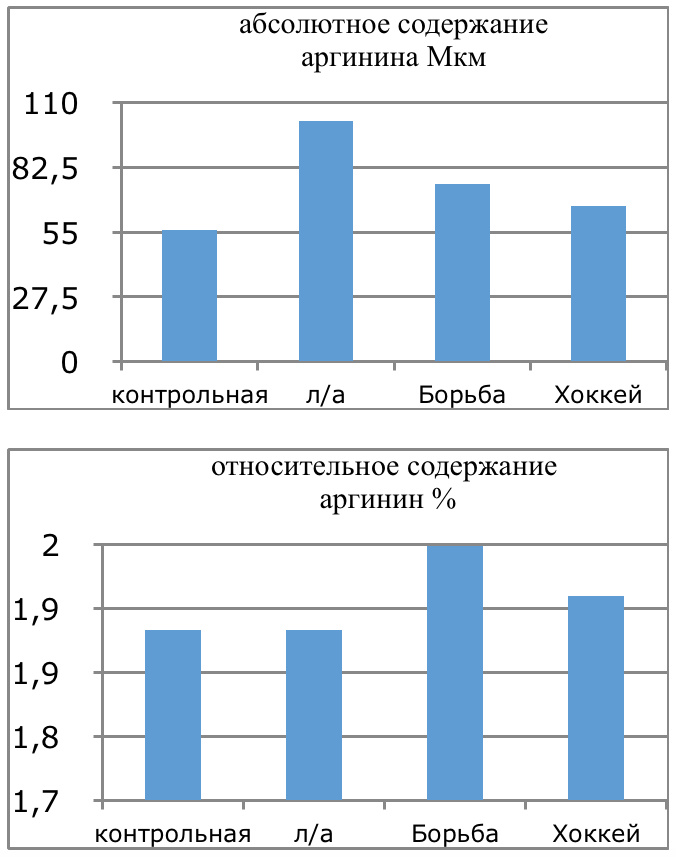
Аргинин (Мкм):
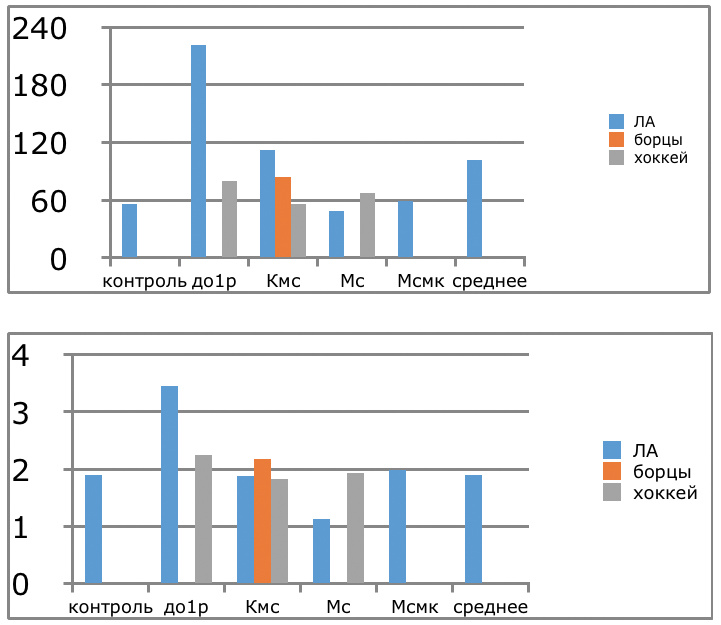
Аргинин (%).
Это метаболит — диаминокислота, образуется в цикле мочевины, преимущественно в печени, но есть ферменты орнитинового цикла и в мышцах и в мозге. Аргинин обладает анаболическим эффектом так как он участвует в образовании полиаминов. Он необходим в образовании креатина. Это нейропротектор [22,24]. Большие количества аргинина образуются в печени, потому особенное значение аргинин имеет для лиц «печенчного» типа -адаптивных механизмов к каковым можно отнести хоккеистов.
Сочетание связей аргинина с психикой говорит о том, что это мощный стабилизатор мембран и борцам, уровень мастерства которых зависит от быстроты реакций и высокой переключаемости нейронов мозга, он в больших количествах не нужен (Табл.4.19).
Таблица 4. 19 Корреляционные связи у борцов, ЛА и хоккеистов (БОРЦЫ, ЛЕГКОАТЛЕТЫ, ХОККЕИСТЫ):
| Показатели | арг(относит. К-во) | арг(абсолют. К-во) | арг(относит. К-во) | Арг(абсолют. К-во) | Арг(относит. К-во) | Арг(абсолют. К-во) |
| отнширтаза | -,695* | |||||
| отндлру | ,828* | |||||
| окрголов | -,917* | |||||
| соотнголов | ,833* | ,757* | ||||
| кум | .c | .c | 1,000** | |||
| санактив | ,998* | ,999* | ||||
| саннастр | -,998* | |||||
| СКР | 1,000** | 1,000** | ,957* | |||
| ПМР | 1,000** | 1,000** | ,955* | |||
| торзсенс | ,917* | -1,000** | -1,000** | ,993** | ||
| ToeA | ,837* | ,812* | ||||
| NISP | -,659* | 1,000** | -1,000** | |||
| SV_ | ,766* | -1,000** | -1,000** | |||
| ** Корреляция значима на уровне 0.01 (2-сторон.) | ||||||
| * Корреляция значима на уровне 0.05 (2-сторон.). | ||||||
ОБЩЕЕ КОЛИЧЕСТВО:
| аргинин | борцы | ЛА | хоккей | |||
| коррел | В т.ч. отр | коррел | В т.ч. отр | коррел | В т.ч.отр | |
| Морфометрия | 1 | 1 | 2 | 1 | 3 | 1 |
| Кардиореспираторная система | 2 | 1 | 4 | 3 | ||
| Психологические особенности | 1 | 5 | 4 | 1 | ||
| Всего 22 | 4 | 2 | 7 | 1 | 11 | 5 |
ВСЕГО КОРРЕЛЯЦИОННЫХ СВЯЗЕЙ:
|
БОРЦЫ |
ЛА |
ХОККЕИСТЫ |
||||
| ВСЕГО | 124 | 71 | 124 | 49 | 111 | 56 |
| морфометрия | 52 | 29 | 75 | 19 | 49 | 22 |
| Кардиореспираторная система | 44 | 25 | 18 | 15 | 35 | 22 |
| Психологические особенности | 30 | 17 | 31 | 15 | 27 | 11 |
| Положит.-отрицат. | (+) | (-) | (+) | (-) | (+) | (-) |
ОБЩЕЕ ЗАКЛЮЧЕНИЕ
На основании наших предшествующих исследований и данных литературы можно с определенностью утверждать, что в особенностях ответа организма на стрессовое воздействие в значительной степени влияют особенности его азотистого обмена, в частности содержание в тканях свободных аминокислот и их соотношения. Нами даже было выдвинуто предположение, что по особенностям обмена аминокислот и преимущественному их образованию в мозге или печени условно можно выделить «МОЗГОВЫЕ» и «ПЕЧЕНОЧНЫЕ» адаптивные типы, совершенно по-разному реагирующие на стрессовые воздействия [197].
Для уточнения роли аминокислот в адаптивных механизмах животных и человека были проведены исследования аминокислотного состава тканей на морских свинках и крысах — животных, имеющих принципиально отличные механизмы адаптации к воздействию повреждающих факторов.
Для того чтобы проверить насколько сопоставимы полученные результаты на животных с изменениями количества аминокислот при стрессовом воздействии у человека проведены серии исследований на спортсменах –профессионалах.
Физическая нагрузка является одним из наиболее мощных и адекватных стрессовых воздействий на животных и человека. Но в разных видах спорта эта нагрузка различна и кроме того сочетается с различной степенью психологической напряженности. То есть получается двойное стрессовое воздействие различной выраженности. Вполне естественно, что переносимость таких воздействий будет зависит от особенностей адаптивных возможностей организма и его способности совершенствовать эти системы в процессе тренировок и соревнований.
При исследовании особенностей изменений количества аминокислот у животных под воздействием стрессовых повреждающих факторов установлено, что большое значение в устойчивости к этим воздействиям имеет обеспеченность организма аминокислотами- адаптогенами, участвующими в биосинтезе гормонов и других биологически активных веществ, антиоксидантных механизмах, повышающих устойчивость мембран к повреждающим воздействиям, детоксикационных процессах и ряде других важных механизмах адаптации.
Так крысы синтезирующие в печени огромные количества таурина (антиоксиданта, стабилизатора мембран, детоксицирующего вещества) и аскорбиновой кислоты оказались гораздо более устойчивы к токсическим воздействиям, кровопотере, заражению МБТ, чем морские свинки. Морские свинки не способны синтезировать аскорбиновую кислоту (так же как и человек) и ориентируются в своих защитных механизмах на систему глутатиона-трипептида, состоящего из глутаминовой кислоты, цистеина и глицина. Соответственно в их тканях имеется повышенное количество этих аминокислот и высокоактивная система глутатиона, участвующая в антиоксидантных, детоксикационных механизмах, транспорте аминокислот и других метаболитов в клетку. Кроме того, что морские свинки не способны синтезировать аскорбиновую кислоту, в их тканях содержится очень малое количество таурина. Все эти различия определяют то обстоятельство, что морские свинки гораздо менее устойчивы к токсическим воздействиям, заражению МБТ, кровопотере.
Но особенности защитных механизмов морской свинки с преобладанием в них системы глутатиона делают ее гораздо более устойчивой по отношению к мутагенным воздействиям и эмоциональному стрессу, чем крысы. При жестких экстремальных воздействиях в тканях генерируются повышенное количество свободных радикалов кислорода, разрушающих мембраны клеток, усиливается апоптоз к чему более устойчивы защитные адаптивные системы крысы. А вот при длительных но более мягких, например, мутагненных воздействиях ксенобиотиков или при хроническом эмоциональном стрессе в защитных механизмах ведущее значение имеет система глутатиона, что и делает морскую свинку более устойчивой к ним , чем крысу.
Если бы нам удалось обнаружить подобные различия роли аминокислот – адаптогенов в механизмах адаптации к стрессовым воздействиям у спортсменов в различных видах спорта и на различном уровне мастерства, то это существенно прояснило роль аминокислот в устойчивости организма к физическим нагрузкам. Это позволило бы применять их более целенаправленно в различных видах спорта для повышения переносимости спортивных нагрузок.
Из полученных нами результатов следует, что аминокислоты играют существенную роль в формировании адаптивных механизмов у спортсменов- профессионалов.
Так уже по анализу общего содержания аминокислот в плазме крови видно, что их количество значительно выше у легкоатлетов, чем у борцов и хоккеистов. Одновременно у них наблюдается более высокий уровень глюкокортикоидов. Нормализация уровня этих стрессовых гормонов происходит только у МСМК.
У борцов и хоккеистов такие сдвиги наблюдаются только у перворазрядников, то есть когда еще нет профессиональной адаптированности организма и имеется хроническая стрессовая реакция.
Обращает на себя внимание то, что у всех исследуемых групп спортсменов в разных видах спорта происходят разнонаправленные изменение количества аминокислот крови по сравнению с контролем, так и мере роста их профессионализма. Но при этом имеются как «неспецифические» аминокислоты которые коррелируют с изменениями морфометрических параметров у всех спортсменов, например, аспарагиновая кислота, глицин, гистидин, орнитин, аргинин. В то же время имеются и «специфические» аминокислоты, динамика содержания которых характерна только для определенных видов спорта. Например, для борцов это глутаминовая кислота и цистеин; для легкоатлетов это аспартат,треонин, серин, глутамин, аланин; для хоккеистов-метионин, таурин, глутамат.
По показателям корреляции аминокислот плазмы крови и данных импедансного исследования кардиореспираторной системы «неспецифических» аминокислот практически нет. При этом у борцов почти все аминокислоты в той или иной степени принимают участие в регуляторных механизмах совершенствования адаптивных механизмов этой системы. Среди которых выделяются количеством коррелятивных связей аспартат, треонин, цистеин, орнитин.
У легкоатлетов только 4 аминокислоты влияют на физиологические показатели, среди которых отчетливо доминирует триптофан. У хоккеистов в процессах регуляции этих систем принимает участие глицин, глутамат, глутамин, пролин, цистеин.
Что касается регуляции психологического состояния то здесь так же имеются группа «неспецифических» аминокислот характерная для всех спортсменов: глицин, аланин, валин, лейцин, изолейцин, тирозин. Специфика же в регуляции этого уровня адаптивных механизмов у борцов принадлежит глутамату, глутамину, триптофану; у легкоатлетов — пролину и аргинину; у хоккеистов — таурину.
Очевидно различия в доминировании определенных аминокислот адаптогенов у спортсменов различных спецификаций определяют различный характер адаптивных процессов по мере роста мастерства.
Мощный адаптоген таурин образующийся в печени и преобладающий у хоккеистов делает их адаптивный тип похожим на адаптивный тип крысы («ПЕЧЕНОЧНЫЙ АДАПТИВНЫЙ ТИП»). Очевидно поэтому они гораздо более устойчивы к экстремальным стрессовым воздействиям, чем борцы и легкоатлеты. Период их адаптации к спортивным нагрузка гораздо короче, чем у последних. До уровня полной адаптированности (автоматизма) легкоатлеты доходят примерно за 5 лет, борцы за 4 года, хоккеисты за 3 (Схема 3).
Схема 3. Особенности хронической стрессовой реакции у спортсменов (по Ф.З. Меерзону и М.Г. Пшенниковой):
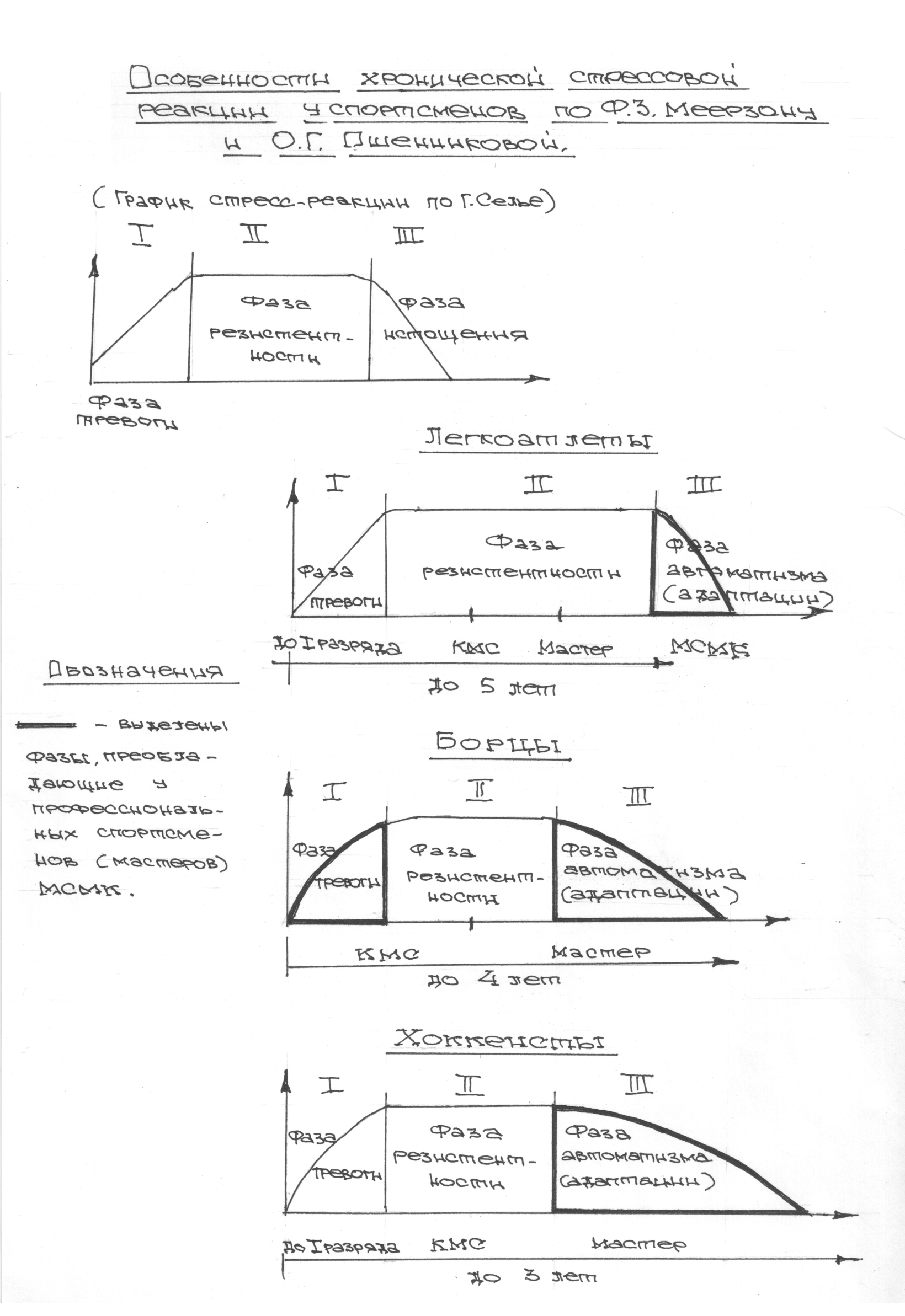
У борцов в адаптивных механизмах преобладают дикарбоновые кислоты, особенно глутамат и его производные, цистеин, то есть это некий промежуточный адаптивный тип: «морская свинка-крыса» или «МОЗГОВИК-ПЕЧЕНОЧНИК». Борцам несколько труднее совершенствовать свой уровень мастерства, чем хоккеистам. Соответственно график совершенствования адаптивных механизмов у них более растянут, чем у хоккеистов. При чем характерно, что у борцов адаптивный ответ на стрессовую реакцию приобретает специфический вид. В нем наличествует как фаза тревоги, так и фаза автоматизма.
У легкоатлетов еще более тонкая настройка адаптивных механизмов. В совершенствовании морфологических структур преобладают аминокислоты в больших количествах присутствующие в нервной и соединительной ткани- аспартат, глиицин, гистидин, аргинин, треонин, серин, глутамин. В регуляции физиологических механизмов у них преобладает триптофан, а психологических — пролин и аргинин. То есть они в еще большей степени в своих адаптивных механизмах похожи на морских свинок(«МОЗГОВИКИ»). И соответственно имеют максимально растянутый период адаптации к спортивным стрессовым нагрузкам, превосходящий таковой и борцов и у хоккеистов.
Полученные результаты могут быть использованы как в целенаправленном использовании аминокислот в спорте, фитнесе, медицине, так и для дальнейших научных исследований, посвященных роли аминокислот в совершенствовании адаптивных механизмов человека к стрессовым воздействиям.
СПИСОК СОКРАЩЕНИЙ
КХА – катехоламины
ГК – глюкокортикоиды
БАВ – биологически-активные вещества
ПАУ – полициклические ароматические углеводороды
ОВП – окислительно-восстановительные процессы
ОСФ – оксигензы смешанных функций
ГМФШ – гиксозомонофосфатный шунт (пентозный цикл)
ФАФС – фосфоаденозин фосфосульфат
УДФГК – уридиндифосфоглюкороновая кислота
ЗАМ – защитно-адаптивные механизмы
АФК – активные формы кислорода
ПОЛ – перекисное окисление липидов
ГГ – глюконеогенез
Г-SH – глутатион восстановленный
Г-SS-Г – глутатион окисленный
ЦНС – центральная нервная система
НЭС – нейроэндокринная система
АТФ – аденозинтрифосфат
АДФ – аденозиндифосфат
АМФ – аденозинмонофосфат
ЦАМФ – цеклический аденазинмонофосфат
ЦГМФ – цеклический гуанозинмонофосфат
ГЭБ – гематоэнцефалитический барьер
ЭС – эмоциональный стресс
АК – аскорбиновая кислота
СТ – соединительная ткань
ЦТК – цикл трикарбоновых кислот (цикл Кребса)
РНК – рибонуклеиновая кислота
ДЦ – дыхательная цепь
Э-М-К – Эмбдена-Мейергофа-Кребса (путь метаболизирования углеводов)
ГОМК – гаммаоксимасляная кислота
ГАМК – гаммааминомасляная кислота
НАДН2 – никотинамидадениндинуклиотид восстановленный
НАДФН2 – никотинамидадениндинуклиотидфосфат восстановленный
ЛПОНП – липопротеиды очень низкой плотности
ЛПНП – липопротеиды низкой плотности
ЛПВП – липопротеиды высокой плотности
СЖК – свободные жирные кислоты
1р. — первый спортивный разряд
КМС — кандидаты в мастера спорта
МС — мастера спорта
МСМК — мастера спорта международного класса
-
— сокращения по специальным методам исследования приведены в приложении.
ОГЛАВЛЕНИЕ
ВВЕДЕНИЕ………………………………………………………………
ГЛАВА 1
-
Роль аминокислот в механизмах адаптации к стрессовым нагрузкам……………………………………………………………
-
Аминокислоты в метаболической «РЕКЕ» филогенеза…………
-
Аминокислоты- адаптогены в механизмах регуляции жизнедеятельности клетки……………………………………………
-
Важнейшие защитно-адаптивные системы организма………………
-
Роль аминокислот- адаптогенов в механизмах реализации стрессовой реакции…………………………………………………….
-
Роль трипептида глутатиона и входящих в него аминокислот в механизмах адаптации……………………………………………………….
-
Мозг и печень как два полюса образования адаптогенов организма. «МОЗГОВИКИ»- «ПЕЧЕНОЧНИКИ»…………………
-
ЭКСПЕРИМЕНТАЛЬНЫЕ ИССЛЕДОВАНИЯ……………………..
-
Роль особенностей метаболизма аминокислот, связанных с обменом соединительной ткани в механизмах адаптации организма к стрессовым воздействиям в опытах на морских свинках и крысах……………………………………
-
Особенности метаболизма аминокислот связанных с образованием фосфолипидов мембран у животных с различной чувствительностью к туберкулезу и мутагенным ксенобиотикам (морские свинки и крысы)………………………….
-
-
Аминокислоты и их производные в формировании «МОЗГОВОГО» И «ПЕЧЕНОЧНОГО» адаптивного типа при стрессовых воздействиях………………………………………………..
-
ГЛАВА 2
-
Особенности обмена аминокислот у спортсменов различных спецификаций……………………………………………………………
-
Введение…………………………………………………………………
2.2Обзор литературы по аминокислотам у спортсменов………………..
2.3 Материалы и методы исследования…………………………………
2.4. Полученные результаты и их обсуждение…………………………
2.4.1. Содержание аминокислот, имеющих достоверные отличия от
контроля и между группами в плазме крови спортсменов……….
2.4.2 Содержание аминокислот, имеющих достоверное отличие от контроля и между группами в плазме крови у спортсменов различных спецификаций
2.4.3. Резюме по 2 главе…………………………………
ГЛАВА 3
-
Механизмы влияния аминокислот на формирование структурного следа адаптации при спортивных нагрузках на основании корреляционного анализа………………………………
ВВЕДЕНИЕ
3.1. Корреляции показателей морфометрии и содержания аминокислот плазмы крови у представителей разных видов спорта…
3.1.1.Корреляции показателей морфометрии с содержанием аминокислот у борцов………………………
3.1.2.Корреляции показателей морфометрии с содержанием аминокислот у легкоатлетов……………
3.1.3.Корреляции показателей морфометрии с содержанием аминокислот у хоккеистов…………………
3.2 Корреляции между аминокислотным составом плазмы крови и показателями гемодинамики по данным импедансного исследованеия……………………
3.2.1. Корреляционные связи между аминокислотным составом плазмы крови и показателями гемодинамики у борцов……………
3.2.2. Корреляционные связи между аминокислотным составом плазмы крови и показателями гемодинамики у легкоатлетов………
3.2.3. Корреляционные связи между аминокислотным составом плазмы крови и показателями гемодинамики у хоккеистов…………
3.3 Корреляционные связи между показателями внешнего дыхания и аминокислотного состава плазмы крови у спортсменов……………
3.3.1.Корреляционные связи между показателями внешнего дыхания и аминокислотным составом плазмы крови у борцов……………………………………
3.3.2.Корреляционные связи между показателями внешнего дыхания и аминокислотным составом плазмы крови у хоккеистов……………………………
3.4 ПСИХОЛОГИЯ Корреляционные связи между аминокислотным составом плазмы крови спортсменов и их психологическими параметрами
3.4.1 Корреляционные связи показателей темперамента и содержанием аминокислот в плазме крови спортсменов…………….
3.4.1.1. БОРЦЫ……………………………………………………
3.4.1.2. ЛЕГКОАТЛЕТЫ…………………………………………
3.4.1.3. ХОККЕИСТЫ…………………………………………
3.4.2 Корреляционные связи показателей самооценки, активности, настроения(САН) и содержанием аминокислот в плазме крови спортсменов………………
3.4.2.1.БОРЦЫ……………………………………………
3.4.2.2. ЛЕГКОАТЛЕТЫ……………………………………..
3.4.2.3. ХОККИСТЫ………………………………………
3.4.3. Корреляционные связи показателей соревновательной устойчивости( по Мильману) с содержанием аминокислот в плазме крови спортсменов…3.4.3.1.БОРЦЫ……………………………………………………….
3.4.3.2.ХОККЕИСТЫ…………………………………………
3.4.4.Корреляционные связи показателей ригидности (по томскому опроснику Залевского-ТОРЗ) с содержанием аминокислот в плазме крови спортсменов
3.4.4.1.БОРЦЫ……………………………………………………….
3.4.4.2.ХОККЕИСТЫ………………………………………………
Резюме по 3 главе………………………………………………
ГЛАВА 4
ПАСПОРТА ОТДЕЛЬНЫХ АМИНОКИСЛОТ, НАИБОЛЕЕ ШИРОКО ПРИМЕНЯЕМЫХ В СПОРТЕ И ФИТНЕСЕ, ПО МАТЕРИАЛАМ НАШИХ ИССЛЕДОВАНИЙ
Таурин………………………………………………………………….
Аспарагиновая кислота………………………………………
Серин…………………………………………………………………..
Глутаминовая кислота…………………………………………
Глутамин………………………………………………………………
Пролин…………………………………………………………………
Глицин………………………………………………………………….
Аланин………………………………………………………………….
Валин…………………………………………………………………….
Цистеин…………………………………………………………………..
Метионин……………………………………………………………
Изолейцин………………………………………………………….
Лейцин……………………………………………………………………
Тирозин…………………………………………………………………
Фенилаланин…………………………………………………
Триптофан………………………………………………………………..
Орнитин………………………………………………………………….
Гистидин…………………………………………………………………
Аргинин………………………………………………………………….
ПРИЛОЖЕНИЕ………………………………………………………….
СПИСОК ЛИТЕРАТУРЫ……………………………………………….
МОРФОМЕТРИЧЕСКИЕ ПОКАЗАТЕЛИ (сантиметры, килограммы):
| ЛЕГКОАТЛЕТЫ | Возраст
лет |
Рост
См. |
Вес
кг |
Вес органов | Индекс пациента |
| Среднее | 19,61 | 178,10 | 68,14 | 382,88 | 51,45 |
| Стд. ошибка среднего | 0,49 | 1,52 | 1,76 | 7,83 | 0,50 |
| Борцы | возраст | рост | вес | Вес органов | Индекс пациента |
| Среднее | 20,61 | 173,09 | 74,56 | 433,71 | 57,12 |
| Стд. ошибка среднего | 0,81 | 1,39 | 2,15 | 9,21 | 0,42 |
| Хоккей | возраст | рост | вес | Вес органов | Индекс пациента |
| Среднее | 23,51 | 181,27 | 76,81 | 423,40 | 52,12 |
| Стд. ошибка среднего | 0,74 | 0,92 | 1,57 | 7,88 | 0,50 |
| ЛЕГКОАТЛЕТЫ | Длина туловища
См. |
Ширина плеч
См. |
Ширина таза
см |
Длина рук
см |
Длина ног
см |
| Среднее | 53,13 | 39,53 | 28,85 | 77,21 | 96,95 |
| Стд. ошибка среднего | 0,57 | 0,45 | 0,43 | 0,80 | 1,02 |
| Борьба | Длина туловища | Ширина плеч | Ширина таза | Длина руки | Длина ног |
| Среднее | 53,60 | 40,27 | 28,46 | 74,97 | 91,70 |
| Стд. ошибка среднего | 0,73 | 0,47 | 0,39 | 1,06 | 0,96 |
| хоккей | Длина туловища | Ширина плеч | Ширина таза | Длина руки | Длина ног | |
| Среднее | 53,99 | 40,73 | 28,75 | 78,22 | 98,10 | |
| Стд. ошибка среднего | 0,62 | 0,33 | 0,28 | 0,58 | 0,71 | |
| Борцы | Относительная длина туловища
см |
Относительная ширина плеч
см |
Относительная ширина таза
см |
Относительная длина рук
см |
Относительная длина ног
см |
|
| Среднее | 30,9794 | 23,2625 | 16,4376 | 43,3030 | 52,9801 | |
| Стд. ошибка среднего | ,32357 | ,17392 | ,15056 | ,45752 | ,35181 | |
| ЛЕГКОАТЛЕТЫ | Относительная длина туловища | Относительная ширина плеч | Относительная ширин таза | Относительная длина рук | Относительная длина ног | |||
| Среднее | 29,84 | 22,20 | 16,17 | 43,35 | 54,42 | |||
| Стд. ошибка среднего | 0,24 | 0,21 | 0,23 | 0,22 | 0,24 | |||
| ЛЕГКОАТЛЕТЫ | Вес жира
кг |
Вес мышц
кг |
Вес костей
кг |
|||||
| Среднее | 8,03 | 33,79 | 10,75 | |||||
| Стд. ошибка среднего | 0,55 | 1,06 | 0,31 | |||||
| Борцы | Вес жира | Вес мышц | вескостей |
| Среднее | 8,87 | 37,73 | 11,44 |
| Стд. ошибка среднего | 0,55 | 1,21 | 0,34 |
| Хоккей | Вес жира | Вес мышц | вескостей | |
| Среднее | 9,77 | 36,96 | 11,96 | |
| Стд. ошибка среднего | 0,62 | 0,79 | 0,23 | |
| ЛЕГКОАТЛЕТЫ | Относительность жира | Относительность мышц | Относительность костей |
| Среднее | 11,92 | 49,39 | 15,65 |
| Стд. ошибка среднего | 0,88 | 0,64 | 0,21 |
| Борцы | Относительность жира | Относительность мышц | Относительность костей | |
| Среднее | 11,70 | 50,05 | 15,26 | |
| Стд. ошибка среднего | 0,57 | 0,56 | 0,33 | |
| Хоккей | Относительность жира | Относительность мышц | Относительность костей | |
| Среднее | 12,55 | 48,09 | 15,72 | |
| Стд. ошибка среднего | 0,62 | 0,35 | 0,22 | |
|
БОРЦЫ –корреляции с относительным содержанием аминокислот |
асп |
глу |
про |
гли |
ала |
цис |
три |
гис |
арг |
|
возраст |
,884** |
-,829** |
|||||||
|
рост |
-,723* |
||||||||
|
Вес органов |
,682* |
||||||||
|
Индекс прапорциональноси |
,749* |
||||||||
|
Отн. Шир. плеч |
,687* |
||||||||
|
Шир. таза |
-,668* |
-,790* |
-,710* |
||||||
|
Отн. Шир. таза |
-,731* |
-,712* |
|||||||
|
Отн. Длина рук |
-,770* |
||||||||
|
Длина ног |
,755* |
,684* |
-,669* |
||||||
|
Вес жира |
,866** |
||||||||
|
Отн. жира |
,723* |
||||||||
|
Отн. мышц |
,836** |
||||||||
|
Отн. костей |
-,852** |
,744* |
|||||||
|
Отн. орг |
-,727* |
,734* |
|||||||
|
Окр. голов |
-,834** |
-,684* |
|||||||
|
Межтеменное расстояние |
-,721* |
-,677* |
-,773* |
||||||
|
* Корреляция значима на уровне 0.05 (2-сторон.). |
|||||||||
|
** Корреляция значима на уровне 0.01 (2-сторон.). |
|||||||||
|
БОРЦЫ- корреляции с абсолютным содержанием аминокислот |
тау |
асп |
глу |
про |
гли |
ала |
три |
орн |
гис |
арг |
|
возраст |
,786** |
-,705* |
||||||||
|
рост |
,712* |
|||||||||
|
Вес орг. |
,720* |
|||||||||
|
Индекс прапорциональности |
-,685* |
-,744* |
||||||||
|
Шир плеч |
-,750* |
|||||||||
|
Отн. Шир. плеч |
-,742* |
|||||||||
|
Шир. таза |
,723* |
,681* |
||||||||
|
отн .шир таза |
-,695* |
|||||||||
|
Длин. руки |
,691* |
-,767* |
||||||||
|
Отн. Дина рук |
-,718* |
-,776* |
||||||||
|
Длина ног |
,893** |
,692* |
||||||||
|
Вес жира |
,742* |
-,676* |
||||||||
|
Вес мышц |
,678* |
|||||||||
|
Отн. мышц |
,764* |
|||||||||
|
Отн. костей |
-,668* |
,738* |
||||||||
|
Окружность головы |
-,679* |
|||||||||
|
ЛЕГКОАТЛЕТЫ – корреляции с относительным содержанием аминокислот |
тау |
асп |
тре |
глу |
глн |
гли |
ала |
мет |
лей |
тир |
фен |
три |
орн |
лиз |
гис |
арг |
||||||||||||||
|
возраст |
,840* |
,928** |
-,813* |
|||||||||||||||||||||||||||
|
рост |
,816* |
-,822* |
||||||||||||||||||||||||||||
|
Вес |
,913* |
|||||||||||||||||||||||||||||
|
Вес органов |
||||||||||||||||||||||||||||||
|
Индекс прапорциональности |
,833* |
|||||||||||||||||||||||||||||
|
Длина туловища |
||||||||||||||||||||||||||||||
|
Отн. Длина туловища |
,859* |
|||||||||||||||||||||||||||||
|
Шир. плеч |
||||||||||||||||||||||||||||||
|
Отн. Шир. плеч |
-,847* |
|||||||||||||||||||||||||||||
|
Ширина таза |
-,871* |
|||||||||||||||||||||||||||||
|
Отн. Шир. таза |
,896* |
,812* |
||||||||||||||||||||||||||||
|
Длина руки |
||||||||||||||||||||||||||||||
|
Отн. Длина рук |
-,883* |
|||||||||||||||||||||||||||||
|
Длина ног |
-,852* |
|||||||||||||||||||||||||||||
|
Отн. Длина ног |
,976** |
-,910* |
||||||||||||||||||||||||||||
|
Вес жира |
,948* |
|||||||||||||||||||||||||||||
|
Отн. жира |
,899* |
-,919* |
,966** |
|||||||||||||||||||||||||||
|
Вес мышц |
||||||||||||||||||||||||||||||
|
Отн. мышц |
||||||||||||||||||||||||||||||
|
Вес костей |
,819* |
|||||||||||||||||||||||||||||
|
Отн. костей |
,972** |
-,895* |
||||||||||||||||||||||||||||
|
Вес органов |
-,945* |
|||||||||||||||||||||||||||||
|
Отн. органов |
,907* |
|||||||||||||||||||||||||||||
|
Окружность голов |
,927* |
-,917* |
||||||||||||||||||||||||||||
|
Переднезадний размер |
,901* |
,983** |
||||||||||||||||||||||||||||
|
Межтеменное расстояние |
-,966** |
|||||||||||||||||||||||||||||
|
ЛЕГКОАТЛЕТЫ- корреляции с абсолютным содержанием аминокислот |
асп |
тре |
сер |
глу |
про |
гли |
ала |
вал |
мет |
иле |
лей |
тир |
орн |
лиз |
гис |
арг |
||||||||||||||
|
возраст |
|
|
|
|
|
|
|
|
|
|
|
-,867* |
|
-,894* |
|
|
||||||||||||||
|
рост |
|
,965** |
,823* |
|
|
,852* |
,978** |
|
|
|
|
|
|
|
|
|
||||||||||||||
|
Индекс прапорциональности |
-,837* |
-,865* |
-,821* |
-,829* |
|
-,855* |
|
|
|
|
|
|
|
|
|
|
||||||||||||||
|
Длина туловища |
|
,935** |
,860* |
|
|
,900* |
,964** |
|
|
|
|
|
|
|
|
|
||||||||||||||
|
Отн. Длина туловища |
|
|
|
|
|
|
|
-,962** |
|
|
-,825* |
|
|
|
|
|
||||||||||||||
|
Отн. Ширина плеч |
|
|
|
|
|
|
|
|
|
|
|
-,831* |
|
|
|
|
||||||||||||||
|
Ширина таза |
|
|
|
|
|
|
|
,935** |
|
|
,831* |
,872* |
|
|
|
|
||||||||||||||
|
Длина руки |
|
,900* |
|
|
|
,845* |
,866* |
|
|
|
|
|
|
|
,821* |
|
||||||||||||||
|
Отн. Длина. рук |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
,891* |
,828* |
||||||||||||||
|
Длина ног |
,854* |
,934** |
,887* |
|
|
,874* |
,950** |
|
|
|
|
|
|
|
|
|
||||||||||||||
|
Отн. Длина ног |
,977** |
|
,871* |
|
|
|
|
|
|
,824* |
|
|
|
|
|
|
||||||||||||||
|
Вес костей |
|
,930** |
|
|
|
|
,941** |
|
|
|
|
|
|
|
|
|
||||||||||||||
|
Отн. костей |
,957* |
,920* |
|
|
|
|
,894* |
|
,903* |
|
|
|
|
|
|
|
||||||||||||||
|
Вес органов |
|
|
,924* |
,884* |
|
,995** |
|
|
|
|
|
|
|
|
|
|
||||||||||||||
|
Отн. органов |
|
|
|
|
,889* |
|
|
|
|
|
|
|
,912* |
|
,881* |
|
||||||||||||||
|
Межтеменное расслояние |
|
|
|
|
|
|
|
|
,932* |
|
|
|
|
|
|
|
||||||||||||||
|
ХОККЕИСТЫ- корреляции с относительным содержанием аминокислот |
тау |
асп |
тре |
сер |
асп |
глу |
про |
гли |
ала |
мет |
лей |
тир |
орн |
лиз |
гис |
арг |
|
возраст |
,841** |
-,806* |
||||||||||||||
|
рост |
||||||||||||||||
|
вес |
,762* |
,716* |
||||||||||||||
|
весор |
,802* |
,743* |
||||||||||||||
|
Индекс пациента |
,947** |
|||||||||||||||
|
Длина туловища |
,803* |
|||||||||||||||
|
Отн. Длина туловища |
,878** |
-,707* |
||||||||||||||
|
ширпл |
,709* |
|||||||||||||||
|
отнширпл |
-,718* |
|||||||||||||||
|
ширтаза |
,845** |
-,711* |
||||||||||||||
|
отнширтаза |
,763* |
-,743* |
||||||||||||||
|
длруки |
||||||||||||||||
|
отндлрук |
-,805* |
|||||||||||||||
|
длног |
||||||||||||||||
|
отндлног |
||||||||||||||||
|
весжира |
,769* |
|||||||||||||||
|
отнжира |
||||||||||||||||
|
весмышц |
-,804* |
,888** |
,719* |
|||||||||||||
|
отнмышц |
-,808* |
-,754* |
||||||||||||||
|
вескостей |
,791* |
,761* |
||||||||||||||
|
отнкостей |
-,829* |
,806* |
||||||||||||||
|
весорг |
-,759* |
-,729* |
||||||||||||||
|
отнорг |
,850** |
,753* |
-,777* |
|||||||||||||
|
окрголов |
-,759* |
-,832* |
,802* |
|||||||||||||
|
перзад |
,817* |
,801* |
||||||||||||||
|
ХОККЕИСТЫ- корреляции с абсолютным содержанием аминокислот |
тау |
асп |
асп |
глу |
про |
мет |
орн |
арг |
|
Отн. Длина туловища |
,758* |
|||||||
|
Ширина таза |
-,731* |
|||||||
|
Отн. Ширина таза |
-,822* |
|||||||
|
Отн. Длина рук |
-,798* |
|||||||
|
Вес мышц |
-,748* |
|||||||
|
Отн. мышц |
-,798* |
,764* |
||||||
|
Окружн. голов |
-,798* |
|||||||
|
Перед-зад. размер |
,799* |
|||||||
|
Соотн. голов |
,757* |
|||||||
СРЕДНИЕ ДАННЫЕ ПО ИМПЕДАНСНЫМ ИССЛЕДОВАНИЯМ СЕРДЕЧНО СОСУДИСТОЙ СИСТЕМЫ:
| Легкоатлеты | |
| PI_ индекс пациента | 44.4+_20.1 |
| HR_ ЧСС | 64.2+_8.1 |
| SpO2 напряжение кислорода | 97.8+-0.5 |
| ToeA плетизмография | 54.6+_36.1 |
| NISP АД систолическое | 124.4+_6.9 |
| NIDP АД диастолическое | 73.1+-5.2 |
| @_S_симпатика | 37.1+-17.7 |
| TrxA крупные сосуды | 179.6+-25.8 |
| SV_ ударный объём | 136.3+-5.3 |
| EF_ фракция выброса | 63.9+-2.5 |
| FW_ ЦВД | 22.8+-25.4 |
| СО_ Минутный объём | 10.0+-2.1 |
| CI_минутный индекс | 6.0+-1.2 |
| DO2 Индекс доставки О2 | 734.5+-25.5 |
| Борцы | |
| PI_ индекс пациента | 63.0+-19.0 |
| HR_ ЧСС | 62.8+-10.1 |
| SpO2 напряжение кислорода | 97.8+-0.8 |
| ToeA плетизмография | 87.9+-54.1 |
| NISP АД систолическое | 132.3+-13.2 |
| NIDP АД диастолическое | 76.6+-7.2 |
| @_S_симпатика | 42.7+-13.7 |
| TrxA крупные сосуды | 150.3+-34.0 |
| SV_ ударный объём | 127.8+-68.9 |
| EF_ фракция выброса | 65.2+-2.3 |
| FW_ ЦВД | 11.8+-5.4 |
| СО_ Минутный объём | 9.3+-5.5 |
| CI_минутный индекс | 5.0+-2.8 |
| DO2 Индекс доставки О2 | 711.2+-306.3 |
| Хоккеисты | |
| PI_ индекс пациента | 44.4+-29.5 |
| HR_ ЧСС | 62.4+-18.4 |
| SpO2 напряжение кислорода | 97.6+-1.3 |
| ToeA плетизмография | 40.4+-39.1 |
| NISP АД систолическое | 129.2+-10.3 |
| NIDP АД диастолическое | 76.2+-5.2 |
| @_S_ симпатика | 35.3+-15.2 |
| TrxA крупные сосуды | 202.2+-57.9 |
| SV_ ударный объём | 130.9+-74.9 |
| EF_ фракция выброса | 65.0+-3.2 |
| FW_ ЦВД | 11.1+-7.6 |
| СО_ Минутный объём | 10.4+-6.1 |
| CI_минутный индекс | 5.7+-3.4 |
| DO2 Индекс доставки О2 | 733.8+-358.0 |
КОРРЕЛЯЦИОННЫЕ СВЯЗИ ПОКАЗАТЕЛЕЙ ИМПЕДАНСНОГО ИССЛЕДОВАНИЯ СЕРДЕЧНО СОСУДИСТОЙ СИСТЕМЫ И СВОБОДНЫХ АМИНОКИСЛОТ ПЛАЗМЫ КРОВИ:
|
БОРЦЫ |
||||||||||||||
|
Корреляции с абсолютным содержанием аминокислот |
асп |
тре |
сер |
глу |
гли |
ала |
цис |
лей |
тир |
фен |
три |
лиз |
гис |
арг |
|
PI_ |
,846** |
,744* |
,644* |
|||||||||||
|
HR_ |
||||||||||||||
|
SpO2 |
-,640* |
|||||||||||||
|
ToeA |
||||||||||||||
|
NISP |
,632* |
-,718* |
-,863** |
-,796** |
-,634* |
-,742* |
-,764* |
-,651* |
-,882** |
-,659* |
||||
|
NIDP |
||||||||||||||
|
@__S_ |
||||||||||||||
|
TrxA |
||||||||||||||
|
SV_ |
,766* |
|||||||||||||
|
EF_ |
||||||||||||||
|
Fw_ |
-,868** |
|||||||||||||
|
CO_ |
,659* |
,661* |
||||||||||||
|
CI_ |
,689* |
,679* |
||||||||||||
|
DO2 |
,746* |
,662* |
,705* |
|||||||||||
|
* Корреляция значима на уровне 0.05 (2-сторон.). |
||||||||||||||
|
** Корреляция значима на уровне 0.01 (2-сторон.). |
||||||||||||||
|
Корреляции с относительным содержанием аминокислот |
асп |
тре |
глу |
гли |
вал |
цис |
мет |
лей |
тир |
орн |
|
PI_ |
,682* |
-,739* |
-,749* |
-,635* |
||||||
|
HR_ |
-,833** |
|||||||||
|
SpO2 |
,635* |
|||||||||
|
ToeA |
||||||||||
|
NISP |
,826** |
|||||||||
|
NIDP |
-,767** |
-,787** |
||||||||
|
@__S_ |
,715* |
|||||||||
|
TrxA |
||||||||||
|
SV_ |
||||||||||
|
EF_ |
-,659* |
|||||||||
|
Fw_ |
-,745* |
,891** |
,761* |
|||||||
|
CO_ |
-,640* |
-,740* |
||||||||
|
CI_ |
-,706* |
|||||||||
|
DO2 |
-,669* |
|||||||||
|
* Корреляция значима на уровне 0.05 (2-сторон.). |
||||||||||
|
** Корреляция значима на уровне 0.01 (2-сторон.). |
||||||||||
|
ЛЕГКОАТЛЕТЫ- корреляции с абсолютным содержанием аминокислот |
тау |
асп |
сер |
цис |
мет |
иле |
фен |
три |
|
PI_ |
-,939* |
-,891* |
||||||
|
HR_ |
,897* |
|||||||
|
SpO2 |
-,919* |
|||||||
|
ToeA |
,884* |
,935* |
||||||
|
NISP |
,670* |
,735* |
||||||
|
NIDP |
||||||||
|
@__S_ |
-,890* |
-,879* |
||||||
|
TrxA |
||||||||
|
SV_ |
||||||||
|
EF_ |
-,882* |
|||||||
|
Fw_ |
,889* |
|||||||
|
CO_ |
||||||||
|
CI_ |
||||||||
|
DO2 |
||||||||
|
* Корреляция значима на уровне 0.05 (2-сторон.). |
||||||||
|
** Корреляция значима на уровне 0.01 (2-сторон.). |
||||||||
|
ЛЕГКОАТЛЕТЫ – корреляции с относительным содержанием аминокислот |
Тау |
асп |
сер |
глу |
фен |
три |
лиз |
гис |
|
PI_ |
-,956* |
-,882* |
||||||
|
HR_ |
||||||||
|
SpO2 |
-,945* |
|||||||
|
ToeA |
-,932* |
|||||||
|
NISP |
-,810* |
|||||||
|
NIDP |
||||||||
|
@__S_ |
-,928* |
|||||||
|
TrxA |
||||||||
|
SV_ |
-,909* |
|||||||
|
EF_ |
-,884* |
-,993** |
,884* |
|||||
|
Fw_ |
-,919* |
|||||||
|
CO_ |
||||||||
|
CI_ |
||||||||
|
DO2 |
-,906* |
|||||||
|
** Корреляция значима на уровне 0.01 (2-сторон.). |
||||||||
|
* Корреляция значима на уровне 0.05 (2-сторон.). |
||||||||
|
ХОККЕИСТЫ- корреляци с абсолютным содержанием аминокислот |
тре |
сер |
асп |
глу |
глн |
гли |
цис |
лей |
фен |
лиз |
арг |
|
PI_ |
|||||||||||
|
HR_ |
|||||||||||
|
SpO2 |
,915* |
,980** |
,924** |
,863* |
|||||||
|
ToeA |
,812* |
||||||||||
|
NISP |
|||||||||||
|
NIDP |
-,940* |
||||||||||
|
@__S_ |
-,911* |
-,934** |
-,915* |
-,814* |
-,838* |
-,814* |
-,859* |
||||
|
TrxA |
,848* |
,824* |
|||||||||
|
SV_ |
1,0** |
1,0** |
1,0** |
-1,0** |
1,00** |
1,0** |
1,0** |
-1,0** |
1,00** |
1,0** |
-1,0** |
|
EF_ |
|||||||||||
|
Fw_ |
|||||||||||
|
CO_ |
-,911* |
||||||||||
|
CI_ |
-,900* |
||||||||||
|
DO2 |
-,909* |
||||||||||
|
* Корреляция значима на уровне 0.05 (2-сторон.). |
|||||||||||
|
** Корреляция значима на уровне 0.01 (2-сторон.). |
|||||||||||
|
ХОККЕИСТЫ- корреляции с относительным содержанием аминокислот |
тре |
асп |
глу |
вал |
мет |
три |
гис |
арг |
|
PI_ |
,893* |
|||||||
|
HR_ |
-,815* |
|||||||
|
SpO2 |
,889* |
|||||||
|
ToeA |
,906* |
,837* |
||||||
|
NISP |
-,906* |
-,923* |
||||||
|
NIDP |
-,936* |
|||||||
|
@__S_ |
||||||||
|
TrxA |
-,839* |
|||||||
|
SV_ |
1,000** |
1,000** |
1,000** |
-1,000** |
1,000** |
1,000** |
-1,000** |
|
|
EF_ |
||||||||
|
Fw_ |
||||||||
|
CO_ |
||||||||
|
CI_ |
||||||||
|
DO2 |
||||||||
|
* Корреляция значима на уровне 0.05 (2-сторон.). |
||||||||
|
** Корреляция значима на уровне 0.01 (2-сторон.). |
||||||||
ПОКАЗАТЕЛИ РЕСПИРАТОРНОЙ СИСТЕМЫ (литры):
| ЛЕГКОАТЛЕТЫ | Резервный объем вдоха | Резервный объем выдоха | Жизненная емкость фактическая | Жизненная емкость расчетная
литры |
Дыхательный Объем максимальная
литры |
| Среднее | 1,70 | 2,67 | 5,19 | 5,35 | 2,57 |
| Стд. ошибка среднего | 0,80 | 0,88 | 0,23 | 0,05 | 0,37 |
| Борцы | Резервный объем вдоха | Резервный объем выдоха | Жизненная емкость фактическая | Жизненная емкость расчетная | Дыхательный Объем максимальная |
| Среднее | 1,25 | 3,07 | 5,36 | 4,84 | 3,22 |
| Стд. ошибка среднего | 0,17 | 0,30 | 0,24 | 0,07 | 0,14 |
| Хоккеисты | Резервный объем вдоха | Резервный объем выдоха | Жизненная емкость фактическая | Жизненная емкость расчетная | Дыхательный Объем максимальная | |
| Среднее | 1,55 | 3,13 | 5,90 | 5,29 | 3,47 | |
| Стд. ошибка среднего | 0,12 | 0,26 | 0,17 | 0,06 | 0,13 | |
| ЛЕГКОАТЛЕТЫ | Минутный Объем Дыхательный максимальная | Удельный Дыхательный Объем максимальная | пвц170 | Пвц удельная | Критическая удельная мощность |
| Среднее | 90,25 | 6,60 | 1068,63 | 16,10 | 2,92 |
| Стд. ошибка среднего | 19,75 | 2,60 | 289,27 | 4,70 | 0,28 |
| Борцы | Минутный Объем Дыхательный максимальная | Удельный Дыхательный Объем максимальная | пвц170 | Пвц удельная | Критическая удельная мощность |
| Среднее | 155,17 | 7,96 | 1348,76 | 18,35 | 3,14 |
| Стд. ошибка среднего | 31,03 | 1,38 | 71,77 | 1,03 | 0,13 |
| Хоккей | Минутный Объем Дыхательный максимальная | Удельный Дыхательный Объем максимальная | пвц170 | Пвц удельная | Критическая удельная мощность |
| Среднее | 150,38 | 6,46 | 1690,24 | 21,67 | 3,12 |
| Стд. ошибка среднего | 16,24 | 0,71 | 71,84 | 0,86 | 0,09 |
КОРРЕЛЯЦИОННЫЕ СВЯЗИ МЕЖДУ ПОКАЗАТЕЛЯМИ РЕСПИРАТОРНОЙ СИСТЕМЫ СПОРТСМЕНОВ И СВОБОДНЫМИ АМИНОКИСЛОТАМИ ПЛАЗМЫ КРОВИ:
|
БОРЦЫ-корреляции с относительным содержанием аминокислот |
асп |
глн |
ала |
иле |
лей |
три |
лиз |
|
ровд |
-,722* |
||||||
|
ровыд |
-,719* |
,760* |
,840** |
,762* |
|||
|
жэлфакт |
-,786* |
||||||
|
жэлрасч |
|||||||
|
ДОмакс |
,746* |
||||||
|
МОДмакс |
-1,000** |
1,000** |
1,000** |
1,000** |
-1,000** |
-1,000** |
|
|
удДОмакс |
-1,000** |
1,000** |
1,000** |
1,000** |
-1,000** |
-1,000** |
|
|
пвц170 |
|||||||
|
пвцудель |
,707* |
-,734* |
|||||
|
кум |
-,785* |
|
БОРЦЫ-корреляции с абсолютным содержанием аминокислот |
асп |
тре |
сер |
асп |
глу |
гли |
цис |
три |
орн |
|
ровд |
,760* |
,793* |
,830* |
||||||
|
ровыд |
-,782* |
-,780* |
-,744* |
-,747* |
-,761* |
||||
|
жэлфакт |
-,761* |
-,894** |
|||||||
|
жэлрасч |
|||||||||
|
ДОмакс |
-,736* |
-,726* |
|||||||
|
МОДмакс |
1,000** |
1,000** |
1,000** |
.c |
1,000** |
1,000** |
1,000** |
.c |
1,000** |
|
удДОмакс |
1,000** |
1,000** |
1,000** |
.c |
1,000** |
1,000** |
1,000** |
.c |
1,000** |
|
пвц170 |
|||||||||
|
пвцудель |
,747* |
||||||||
|
кум |
|
ХОККЕИСТЫ- корреляции с относительным содержанием аминокислот |
асп |
тре |
сер |
глу |
гли |
ала |
мет |
тир |
орн |
лиз |
гис |
арг |
|
ровд |
||||||||||||
|
ровыд |
||||||||||||
|
жэлфакт |
-1,000* |
-,999* |
||||||||||
|
жэлрасч |
-1,000** |
1,000* |
||||||||||
|
ДОмакс |
-,999* |
-1,000* |
||||||||||
|
МОДмакс |
-1,000* |
,997* |
-1,000* |
|||||||||
|
удДОмакс |
-,999* |
,999* |
||||||||||
|
пвц170 |
||||||||||||
|
пвцудель |
1,000* |
|||||||||||
|
кум |
,999* |
1,000** |
||||||||||
|
** Корреляция значима на уровне 0.01 (2-сторон.). |
||||||||||||
|
* Корреляция значима на уровне 0.05 (2-сторон.). |
||||||||||||
|
ХОККЕИСТЫ –корреляции с абсолютным содержанием аминокислот |
тау |
асп |
тре |
глу |
гли |
мет |
иле |
тир |
орн |
гис |
арг |
|
ровд |
-,997* |
-,639* |
|||||||||
|
ровыд |
1,000* |
,999* |
|||||||||
|
жэлфакт |
1,000** |
,999* |
|||||||||
|
жэлрасч |
-1,000* |
-,999* |
-,999* |
||||||||
|
ДОмакс |
|||||||||||
|
МОДмакс |
-,998* |
,998* |
|||||||||
|
удДОмакс |
-1,000* |
||||||||||
|
пвц170 |
|||||||||||
|
пвцудель |
,998* |
||||||||||
|
кум |
-,999* |
,999* |
|||||||||
|
** Корреляция значима на уровне 0.01 (2-сторон.). |
|||||||||||
|
* Корреляция значима на уровне 0.05 (2-сторон.). |
|||||||||||
ПОКАЗАТЕЛИ ПСИХОЛОГИЧЕСКОГО СОСТОЯНИЯ СПОРТСМЕНОВ (условные единицы):
| ЛЕГКОАТЛЕТЫ | экстраверсия | нейротизм | САН самочувствие | Сан активность | Сан настроение |
| Среднее | 15,00 | 9,75 | 71,05 | 70,47 | 74,29 |
| Стд. ошибка среднего | 0,47 | 1,12 | 2,27 | 2,37 | 2,53 |
| Борцы | экстраверт | нейротизм | САН самочувствие | Сан активность | Сан настроение |
| Среднее | 13,90 | 10,63 | 67,03 | 63,03 | 70,31 |
| Стд. ошибка среднего | 0,43 | 0,64 | 1,96 | 1,76 | 2,29 |
| Хоккей | Айзенка экстраверт | Айзенка нейротизм | САН самочувствие | Сан активность | Сан настроение |
| Среднее | 15,24 | 10,48 | 68,12 | 64,88 | 73,87 |
| Стд. ошибка среднего | 0,37 | 0,65 | 1,88 | 1,99 | 2,36 |
| Легкоатлеты | Соревновательная эмоциональная устойчивость(СЭУ) | само регуляция
(СР) |
Энергия мотивации
(ЭМ) |
Соревновательная стабильность
(СТ) |
| Среднее | -1,14 | 0,42 | 2,00 | 0,14 |
| Стд. ошибка среднего | 1,14 | 0,89 | 1,02 | 0,85 |
| Борцы | Соревновательная эмоциональная устойчивость | само регуляция | Энергия мотивации | Соревновательная стабильность |
| Среднее | -3,12 | -1,42 | 0,21 | -1,51 |
| Стд. ошибка среднего | 0,43 | 0,59 | 0,40 | 0,39 |
| Хоккей | Соревновательная эмоциональная устойчивость | само регуляция | Энергия мотивации | Соревновательная стабильность |
| Среднее | -1,64 | -0,58 | 2,23 | -0,88 |
| Стд. ошибка среднего | 0,39 | 0,47 | 0,53 | 0,58 |
| ЛЕГКОАТЛЕТЫ | Симптомо комплекс ригидности
(СР) |
Актуальная ригидность(АР) | Преморбидная ригидность
(ПМР) |
Ригидность как состояние
(РС) |
Сенситивная ригидность(СР) | Установочная ригидност(УР) |
| Среднее | 133,37 | 32,18 | 38,56 | 10,43 | 27,06 | 38,06 |
| Стд. ошибка среднего | 4,81 | 2,32 | 1,48 | 1,24 | 3,95 | 2,98 |
КОРРЕЛЯЦИОННЫЕ СВЯЗИ МЕЖДУ ПОКАЗАТЕЛЯМИ ПСИХОЛОГИЧЕСКОГО СОСТОЯНИЯ СПОРТСМЕНОВ И СОДЕРЖАНИЕМ СВОБОДНЫХ АИМНОКИСЛОТ В ИХ ПЛАЗМЕ КРОВИ
Корреляционных связей между абсолютным содержанием аминокислот в плазме крови у борцов и показателями темперамента нет.
|
Борцы -корреляции с абсолютным содержанием аминокислот |
вал |
цис |
орн |
|
экстраверт |
-,844** |
||
|
нейротизм |
,757* |
,822* |
|
|
* Корреляция значима на уровне 0.05 (2-сторон.). |
|||
|
** Корреляция значима на уровне 0.01 (2-сторон.). |
|||
Корреляционных связей между абсолютным содержанием аминокислот в плазме крови у хоккеистов и показателями темперамента нет.
|
Хоккеисты – корреляции с абсолютным содержанием аминокислот |
тау |
вал |
фен |
|
айзэкстрав |
-,976* |
||
|
Айзнейротизм |
-,988* |
-,999** |
|
|
* Корреляция значима на уровне 0.05 (2-сторон.). |
|||
|
** Корреляция значима на уровне 0.01 (2-сторон.). |
|||
|
Борцы -корреляции с относительным содержанием аминокислот |
глу |
глу |
гли |
три |
|
сансамоч |
-,803* |
|||
|
санактив |
||||
|
саннастр |
,809* |
-,772* |
-,779* |
-,767* |
|
* Корреляция значима на уровне 0.05 (2-сторон.). |
||||
|
** Корреляция значима на уровне 0.01 (2-сторон.). |
||||
|
Борцы- корреляции с абсолютным содержанием аминокислот |
три |
|
сансамоч |
-,763* |
|
санактив |
|
|
саннастр |
|
|
** Корреляция значима на уровне 0.01 (2-сторон.). |
|
|
* Корреляция значима на уровне 0.05 (2-сторон.). |
|
|
Легкоатлеты- корреляции с относительным содержанием аминокислот |
мет |
три |
Гис |
|
сансамоч |
,987* |
||
|
санактив |
-,980* |
,987* |
|
|
саннастр |
|||
|
* Корреляция значима на уровне 0.05 (2-сторон.). |
|||
|
** Корреляция значима на уровне 0.01 (2-сторон.). |
|||
| Хоккеисты- корреляции с относительным содержанием аминокислот | тау | асп | вал | фен | гис |
| сансамоч | -,780* | ||||
| санактив | -,878* | ,759* | |||
| саннастр | ,738** | ,873* | ,695* | 0,869 | |
| * Корреляция значима на уровне 0.05 (2-сторон.). | |||||
| ** Корреляция значима на уровне 0.01 (2-сторон.). | |||||
| Хоккеисты- корреляции с абсолютным содержанием аминокислот | Арг | |||||||||
| сансамоч | ||||||||||
| санактив | ||||||||||
| саннастр | -,998* | |||||||||
| ** Корреляция значима на уровне 0.01 (2-сторон.). | ||||||||||
| * Корреляция значима на уровне 0.05 (2-сторон.). | ||||||||||
| Борцы – корреляции с относительным содержанием аминокислот | сер | глу | ала | цис | тир | орн | гис | |||
| сэу | ,824* | -,784* | -,785* | -,811* | ||||||
| ср | -,770* | |||||||||
| мэ | ,730* | -,783* | -,857** | -,749* | ||||||
| стп | ||||||||||
| * Корреляция значима на уровне 0.05 (2-сторон.). | ||||||||||
| ** Корреляция значима на уровне 0.01 (2-сторон.). | ||||||||||
| Борцы- корреляции с абсолютным содержанием аминокислот | тре | глу |
| сэу | ||
| ср | ||
| мэ | ,762* | ,870** |
| стп | ||
| ** Корреляция значима на уровне 0.01 (2-сторон.). | ||
| * Корреляция значима на уровне 0.05 (2-сторон.). | ||
| Хоккеисты- корреляции с абсолютным содержанием аминокислот | иле | орн |
| сэу | ||
| ср | ||
| мэ | -,981* | |
| стп | -,996** | |
| * Корреляция значима на уровне 0.05 (2-сторон.). | ||
| ** Корреляция значима на уровне 0.01 (2-сторон.). | ||
| Хоккеисты – корреляции с относительным содержанием аминокислот | про | гли | вал | иле | лей | тир | фен |
| сэу | ,954* | ,971* | -,977* | ,997** | ,981* | ||
| ср | |||||||
| мэ | -,977* | ||||||
| стп | -,954* | ||||||
| ** Корреляция значима на уровне 0.01 (2-сторон.). | |||||||
| * Корреляция значима на уровне 0.05 (2-сторон.). | |||||||
| Борцы- корреляции с абсолютным содержанием аминокислот | тау | тре |
| СКР | ||
| АР | ||
| ПМР | ||
| РСО | -,833* | |
| торзсенс | -,833* | |
| УР | ||
| * Корреляция значима на уровне 0.05 (2-сторон.). | ||
| ** Корреляция значима на уровне 0.01 (2-сторон.). | ||
|
Борцы- корреляции с относительным содержанием аминокислот |
сер |
глу |
вал |
иле |
лей |
фен |
лиз |
арг |
|
СКР |
||||||||
|
АР |
||||||||
|
ПМР |
||||||||
|
РСО |
-,888* |
-,937** |
,840* |
,858* |
,956** |
,831* |
,901* |
|
|
торзсенс |
-,867* |
,918** |
,917* |
|||||
|
УР |
||||||||
|
* Корреляция значима на уровне 0.05 (2-сторон.). |
||||||||
|
** Корреляция значима на уровне 0.01 (2-сторон.). |
||||||||
|
Хоккеисты-корреляции с относительным содержанием аминокислот |
гли |
ала |
лей |
арг |
|
СКР |
||||
|
АР |
||||
|
ПМР |
-,996** |
,982* |
||
|
РСО |
,985* |
|||
|
торзсенс |
,951* |
|||
|
УР |
-,958* |
,972* |
||
|
** Корреляция значима на уровне 0.01 (2-сторон.). |
||||
|
* Корреляция значима на уровне 0.05 (2-сторон.). |
||||
Корреляционных связей между показателями ТОРЗ и содержанием аминокислот в плазме крови у хоккеистов нет.
СПИСОК ЛИТЕРАТУРЫ