На пути от естества природы к ноосфере.
Екатеринбург
ОГЛАВЛЕНИЕ
АННОТАЦИЯ……………………………………………………………………………..…..3
ВВЕДЕНИЕ……………………………………………………………………………………….4
1. МЕХАНИЗМЫ АДАПТАЦИИ КАК ФАКТОРЫ НЕПРЕРЫВНОГО СОВЕРШЕНСТВОВАНИЯ ЖИВОЙ МАТЕРИИ……………… 10
1.1 МЕТАБОЛИЧЕСКАЯ «РЕКА» ФИЛОГЕНЕЗА……………………………10
1.2 АМИНОКИСЛОТЫ АДАПТОГЕНЫ В МЕХАНИЗМАХ РЕГУЛЯЦИИ ЖИЗНЕДЕЯТЕЛЬНОСТИ КЛЕТКИ…………………………..21
1.3 ВАЖНЕЙШИЕ ЗАЩИТНО-АДАПТИВНЫЕ СИСТЕМЫ ОРГАНИЗМА……..………………………………………………………………………27
1.4 РОЛЬ АМИНОКИСЛОТ — АДАПТОГЕНОВ В МЕХАНИЗМАХ РЕАЛИЗАЦИИ СТРЕСС – РЕАКЦИИ………………………………………………33
1.5 ПРИМЕРЫ ВЛИЯНИЯ АМИНОКИСЛОТ-АДАПТОГЕНОВ НА МЕХАНИЗМЫ СТРЕСС – РЕАКЦИИ…………………………………..…………41
1.6 СТРЕСС ВОЗДЕЙСТВИЕ, КАК ФАКТОР СТИМУЛЯЦИИ И ИНТЕГРАЦИИ ЗАЩИТНО-АДАПТИВНЫХ СИСТЕМ ОРГАНИЗМА, ПОПУЛЯЦИИ И ОБЩЕСТВА……………………………………………………….48
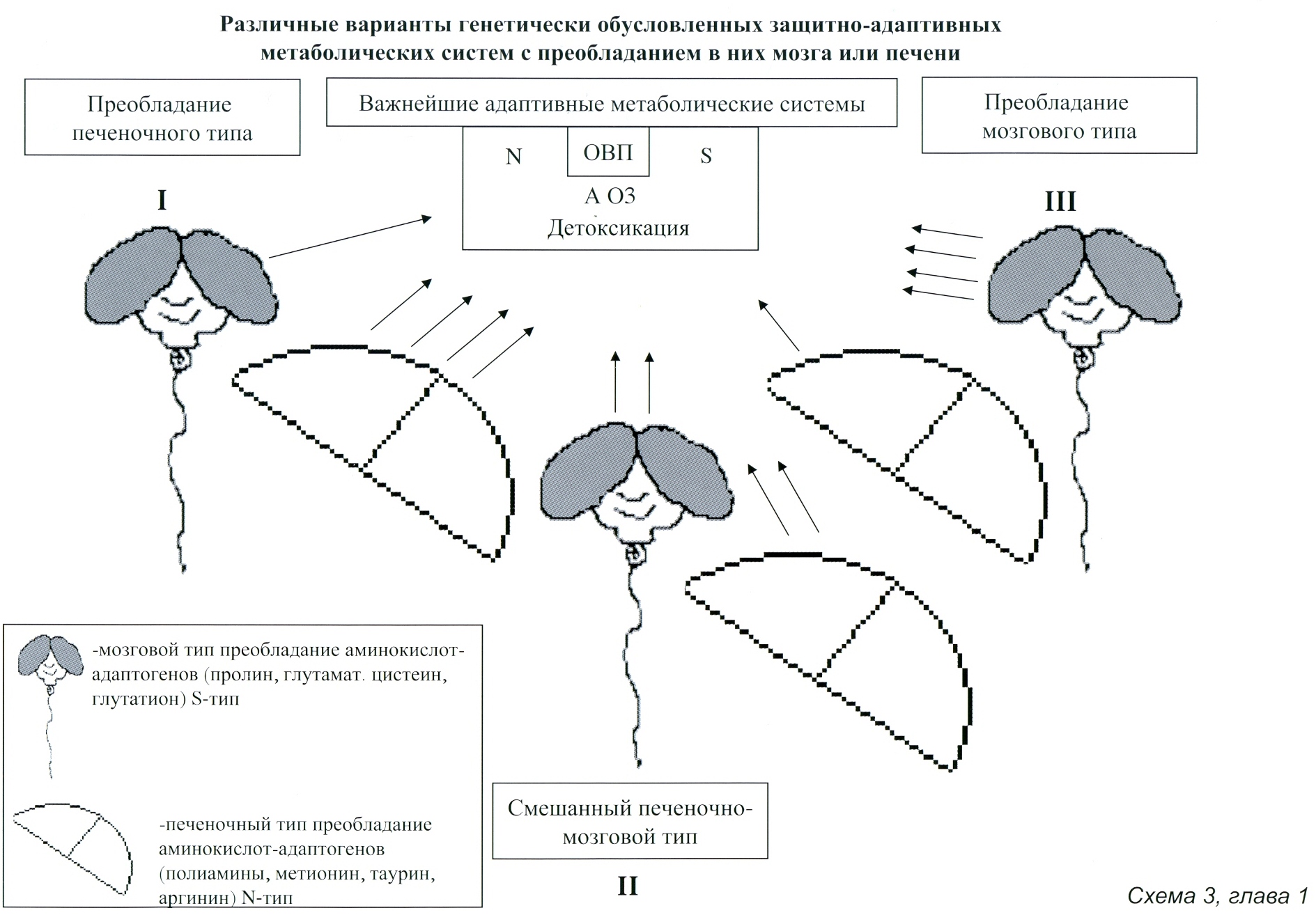
1.7 МОЗГ И ПЕЧЕНЬ, КАК ДВА ОСНОВНЫХ ПОЛЮСА ОБРАЗОВАНИЯ АДАПТОГЕНОВ ОРГАНИЗМА В ЭВОЛЮЦИОННОМ АСПЕКТЕ…………………………………………………………………………………54
2. ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ПСИХОЛОГИЧЕСКОЙ АДАПТИВНОСТИ………………………………………………………………………….75
2.1 ВВЕДЕНИЕ К ЦЕРЕБРАЛЬНОЙ ЭВОЛЮЦИИ КАК ОСНОВЫ АНТРОПОГЕНЕЗА………………………………………………………………………….75
2.2 ЭВОЛЮЦИЯ ДВИЖЕНИЯ………………………………………………………81
2.3 ФИЗИОЛОГИЯ ДВИЖЕНИЯ…………………………………………………….84
2.4 НЕАНДЕРТАЛЕЦ И КРОМАНЬОНЕЦ……………………………………..106
2.5 МАГИЯ………………………………………………………………………………111
2.6 СОЧЕТАННОЕ РАЗВИТИЕ ПРЕДМЕТНОЙ ДЕЯТЕЛЬНОСТИ И ЭМОЦИОНАЛЬНОЙ СИГНАЛЬНОЙ СИСТЕМЫ…………………………….117
2.7 КРОМАНЬОНЦЫ…………………………………………………………………151
3. ПСИХОФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ СОЦИАЛЬНОЙ АДАПТИВНОСТИ…………………………………………………………………………172
3.1 ДИНАМИЗМ СОЦИАЛЬНО-ЭНЕРГИТИЧЕСКИХ УРОВНЕЙ ОБЩЕСТВА…………………………………………………………………………….172
3.2 ТРАНСФОРМАЦИЯ ПСИХОФИЗИОЛОГИЧЕСКИХ АДАПТИВНЫХ ТИПОВ КАК СОЦИАЛЬНО — ЭНЕРГЕТИЧЕСКАЯ ОСНОВА ИСТОРИЧЕСКОГО ПРОЦЕССА……………………………………….179
3.3 ВОЗМОЖНАЯ РОЛЬ И МЕСТО РОССИИ В СОВРЕМЕННОЙ ЦИВИЛИЗАЦИИ…………………………………………………………………………..181
ЛИТЕРАТУРА………………………………………………………………………………186
СПИСОК СОКРАЩЕНИЙ……………………………………………………………….194
Приложение( схемы и рисунки)…………………………………… стр.196 — 205
АННОТАЦИЯ
На основании многолетних научных исследований и анализа литературных источников авторы приходят к выводу, что существующие представления о механизмах адаптации индивидуума и общества можно существенно дополнить и переосмыслить. Найдены объективные критерии оценки адаптивных типов, как животных, так и человека, которые формировались и совершенствовались в процессе развития живой материи. Важнейшее значение в совершенствовании механизмов адаптации имеют мозг и печень. По преобладанию этих органов в механизмах защиты к стрессовым факторам можно условно выделить «мозговиков» и «печеночников» как среди животных, так и среди людей. Метаболические особенности адаптивного типа определяют психофизиологические адаптивные ресурсы индивидуума и влияют на взаимоотношения между индивидуумами. Характер этих отношений формирует определенный социальный организм, развитие которого подчинено объективным законам совершенствования процессов адаптации и гармонизации с окружающим миром.
Полученные результаты позволяют переоценить существующие воззрения на антропогенез и развитие общества. Установлена связь между психологическими и физиологическими сторонами антропогенеза, а так же между метаболизмом отдельных людей и структурой общественных связей. Биологическая основа общества определяет его перспективность с точки зрения социального развития. Современное общество России имеет высокий потенциал социального и культурного развития.
Из рецензии на рукопись Павлова В.А. и соавторов на тему:
«На пути естества природы к ноосфере. Роль России в этом процессе»
В своем оригинальном и обстоятельном труде В.А. Павлов и соавторы рассматривают защитно-адаптивные механизмы живой материи и биоценоза как систему, сложившуюся в результате эволюции Вселенной и жизни на нашей планете, продолжающейся уже более трех миллиардов лет.
Проведенное исследование может представлять как научный, так и научно популярный интерес для преподавателей и студентов, пытающихся с научных позиций (биологии, социологии, экологии, истории, философии) найти ответы на вопросы дальнейшего развития биоценоза, биосферы, человека, цивилизации и роли России в этих процессах.
Новоженов Ю.И. доктор биологических наук, профессор кафедры зоологии Уральского Государственного университета, автор концепции «Культура как форма адаптации».
ВВЕДЕНИЕ
Краткое изложение основ адаптологической концепции развития живой материи и общества.
Известным русским ученым — естествоиспытателем Вернадским В.И., были выдвинуты и в определенной степени обоснованы идеи о том, что неживая природа, развиваясь, дает начало более совершенной по организации и энергопотреблению живой природе. Соприкасаясь — между собой живая и неживая природа дают биосферу — матрицу, на которой живет, развивается и становится все более совершенной живая природа в виде проходящих стадий филогенеза живых организмов. В процессе этого совершенствования повышаются адаптивные возможности организмов. Революционное значение в этом процессе имело появление и совершенствование нервной системы. Вершиной этого эволюционного процесса является становление Ноmо sapiens с его мозгом, способном мыслить, накапливать знания, добытые предшествующими поколениями и за счет этого резко повышать свои адаптивные возможности. В.И. Вернадский сформулировал концепцию, согласно которой развитие живой природы, имеющее характер эволюции, идет от биосферы к ноосфере. Т.е. от царства природы к царству разума, единственно возможной формы развития человечества в условиях современной технократической цивилизации. Апогеем этого процесса, должно стать возникновение ноосферы, когда не природа будет играть человеком и задавать ему законы бытия человек создает гармоничное с природой общество неограниченное в своих возможностях, что собственно соответствует моменту озарения в процессе самопостижения духа по Гегелю.
В противном случае неконтролируемое антропогенное воздействие на природу может необратимо нарушить и подорвать основы существования биосферы на Земле.
Предшественником В.И. Вернадского можно считать великого немецкого философа И.Г. Гердера, рассматривавшего все происходящее на Земле как реализацию высших начал, творящих методом проб и ошибок наиболее совершенные, гармоничные во взаимоотношениях друг с другом формы. Процесс этот направлен от духа Земли, т.е. совершенствования естественных процессов живой природы в ходе эволюции к возникновению и совершенствованию мысли или духа гуманизма в человеческом обществе. Гердер определяет стремление человека к свободе — как стремление к постижению высших сил (законов природы) руководствующих нами.
История человечества — арена превращений духа. Идеи И.Г.Гердера развивает Г.В.Ф. Гегель, который говорит о том, что высшее воплощение духа на Земле — это возникновение великих государств и империй, которые и определяют развитие цивилизации человеческой мысли, духа истории.
Однако к созданию великих государств и империй по силам далеко не всем народам Земли, создавать их могут лишь пассионарныe этносы.
Русский мыслитель, практически наш современник Л.Н. Гумилев разработал концепцию о зарождении и развитии из великих народов пассионарных этносов. Согласно Л.Н. Гумилеву в зарождении таких этносов имеют значение многие факторы биосферы, география и ландшафт где эти народы проживают, их история, контакты с другими народами и еще многое другое.
Однако до конца концепция становления ноосферы В.И. Вернадского и носителей этой ноосферы — пассионарных этносов по Л.Н. Гумилеву так и не были разработаны. Это скорее были пророчества, прозрения, их авторы, базируясь на знаниях, накопленных на начало двадцатого века, не имели возможностей конца двадцатого века, и потому не были в состоянии довести их до уровня науки. Они остались несбывшимися пророчествами. Механизмы происходящего могут быть видны гораздо более полно нашим современникам. Для этого необходимо припасть к истокам человеческой мудрости и интегрировать все наработанное человеческой мыслью за историю развития цивилизации, касающегося как становления человека, так и человеческого общества.
Анализ основ жизнедеятельности и развития живой материи с момента ее зарождения до современного состояния биоценоза с позиций различных научных подходов позволяет утверждать, что основным свойством и одновременно механизмом ее совершенствования является адаптация — приспособление к изменяющимся условиям окружающей среды.
При этом если на ранних этапах зарождения живых существ их совершенствование происходило на уровне обменных процессов, то затем адаптивные механизмы все больше проявляются на уровне функциональном — повышается их активность и плодовитость. По мере остывания планеты возникла необходимость в использовании биоценозом энергии Солнца, что было достигнуто формированием процессов фотосинтеза некоторыми бактериями и растениями. Однако при этом, как побочный продукт фотосинтеза, в атмосфере стал накапливаться кислород — мощный окислитель, который произвел революцию в развитии живой материи. Наиболее приспособленной к использованию кислорода тканью является нервная ткань и особенно головной мозг. Именно их возникновение и совершенствование во многом определило дальнейший процесс эволюции. Этот процесс сопровождается явлением цефализации (постоянного увеличения и совершенствования мозга). Как установлено еще в 19 веке, процессы эволюции, так же как и цефализации, обратного хода не имеют. Они могут затормаживаться, застывать на некоторое время, но затем неизбежно их дальнейшее развитие и совершенствование.
По мере цефализации, в дополнение к явлению физиологической адаптивности, возникло и развивается явление адаптивности психологической. Имея достаточно примитивные формы у животных, такая адаптивность достигла своего апофеоза у приматов и человека. При этом психологическая и физиологическая адаптивность являются как бы двумя сторонами совершенствования процесса адаптации как основного механизма эволюционного процесса живой материи под воздействием изменяющихся факторов окружающей среды.
Во времени этот процесс имеет нарастающий и восходящий характер. Человеческое сознание и человеческая мысль становятся все более мощными и богатыми. Эти явления имеют вполне определенное биологическое основание — головной мозг («инструмент прогресса»).
Американский естествоиспытатель Д.Д. Дана в середине XIX века сформулировал концепцию, согласно которой эволюция животного мира сопряжена с возникновением и совершенствованием центральной нервной системы и мозга. При этом ход эволюции и изменения идут только в сторону все более сложных и совершенных ее систем и никогда наоборот. В дальнейшем было установлено, что нервная ткань и особенно нейроны мозга обладают уникальными биологическими особенностями. Так, обмен головного мозга идет с постоянно высокой интенсивностью. Мозг, составляя 2 % от массы тела человека, потребляет до 25 % кислорода, поглощаемого организмом в покое, при этом необходимо постоянное обеспечение мозга углеводами, расщепление которых дает энергию для его работы. Прекращение поступления кислорода или углеводов даже на несколько минут ведет к потере сознания и гибели нервных клеток, а затем и всего организма. Таким образом, нейроны мозга — это постоянно работающие с высокой интенсивностью сложноорганизованные клетки, контролирующие всю жизнедеятельность организма, его приспособление к изменяющимся условиям окружающей среды — адаптацию.
Нервная ткань имеет существенное отличие от других тканей организма. Так, нейроны, стремятся к максимальному увеличению поверхности за счет появления и роста отростков — дендритов и аксонов и имеют уникальные морфофункциональные особенности позволяющие генерировать разнообразные электрические импульсы. Во многом это определяется строением их мембран и наличием в них нейромедиаторов, обладающих положительными и отрицательными зарядами. Существует симбиотическая концепция происхождения нейронов от микроорганизмов, способных переносить большую кислородную нагрузку, которая подтверждается генетическими особенностями этих клеток и особенно их митохондрий: В функциональном плане это позволяет потреблять нейронам большие количества кислорода и интегрировать химические особенности нейромедиаторов, биологически активных олиго — и полипептидов, образующихся в мозге, с электрофизиологической его функцией (А.М.Хазен, «Разум природы и разум человека» М. 2000).
Это во многом определяет высокую адаптивность нервной ткани и ее главенствующее регуляторное значение.
Так, например, образующиеся в мозге карнозин и анзерин, усиливают работу утомленных мышц, обладая стабилизирующим воздействием на их мембраны и восстанавливающим эффектом на электролитные насосы этих мембран. Т.е., эти вещества обеспечивают функциональные возможности организма в условиях чрезмерных нагрузок в экстремальных ситуациях («второе дыхание»).
Большие количества таурина, цистеина, глутамата, глутамина, аспартата, присутствующие в тканях мозга, являются мощными адаптогенами, значительно повышающими устойчивость организма и разнообразным повреждающим факторам.
Таким образом, мощная ЦНС с большим мозгом — это важнейший регуляторно-метаболический ресурс защитно-адаптивных механизмов, что, очевидно, и определяет существенное превосходство человека над всеми животными в плане адаптивно-приспособительного потенциала организма к меняющимся условиям среды.
По некоторым данным 500-300 млн. лет назад накопление кислорода в атмосфере достигло 40-50%, что сопровождалось интенсивными пожарами древних лесов и последующим снижением концентрации кислорода при накоплении в биосфере большого количества продуктов сгорания органического вещества (каменноугольный период). Образующиеся биологически активные вещества воздействовали на живые организмы, которые были вынуждены приспосабливаться к новым условиям окружающей среды и обезвреживать поступающие извне вредные вещества. Важнейшим органом их обезвреживания стала печень. Сформировался другой важнейший центр адаптации, в котором происходит обезвреживание вредных для организма веществ, как поступающих извне, так и образующихся в ходе обменных процессов, кроме того, здесь стали образовываться биологически активные вещества, влияющие на функцию других адаптивных систем, прежде всего нервной системы и мозга.
Процесс формирования защитно-адаптивных систем и их совершенствование был сопряжен с подбором биологически активных молекул. Так, в ходе эволюции постепенно сформировался универсальный набор метаболитов-регуляторов, обладающих свойствами медиаторов, гормонов, антиоксидантов и других веществ, повышающих адаптивность организма-адаптогенов. Важным фактором является трансформация химических особенностей перечисленных веществ в электрофизиологические сигналы, в наибольшей степени генерируемые мозгом и ЦНС.
На пути совершенствования этих адаптивных систем и органов наибольшего прогресса достигли млекопитающие, среди которых максимальной адаптивности за счет усиления функции печени достигли такие животные как крыса, собаки, лошади и ряд других, а пик цефализации приходится на предшественников человека — троглодитов-неандертальцев, которые, по всей видимости, людьми еще не были. Но их большой мозг (больший по размерам, чем у современного человека) позволял им адаптироваться за счет генерирования особого рода психологической энергии — суггестии (внушения) и психологического подавления окружающего животного мира. Эта данная природой особенность хорошо защищала их от хищников и позволяла им жить как потребители, отбирая добычу у хищников и своих менее суггестивных сородичей, а затем стать их пожирателями-каннибалами. То есть это было абсолютное преобладание «мозгового» психо-адаптивного типа. Однако, суггестируемые для того чтобы выжить, вынуждены были вырабатывать механизмы контрсуггестии — мощные тормозные процессы с участием активного совершенствования механизмов адаптации на уровне печени, контролирующих тормозную функцию мозга. Печень нарабатывает вещества, повышающие устойчивость организма к холоду, интоксикациям, инфекциям, радиации, физической нагрузке и другим повреждающим воздействиям. Очень большое значение имеет наработка печенью вещества, усиливающего тормозные функции мозга – таурина. Именно значительное усиление тормозных процессов, как бы наслаивание их друг на друга в формирующихся молодых структурах мозга, сделали возможным эффективный контроль над процессами психической адаптации и создании мощной защиты от психологической агрессии – суггестии. Одновременно эти же механизмы лежали в основе резкого повышения координации движений, появления зачатков речи и соответственно мышления и способности к вербальным коммуникациям, что предопределило появление первых людей — кроманьонцев. Все это сделало возможным появление нового небывалого ранее в природе адаптивного организма с преобладанием социальной адаптивности — человеческой общины. Так возникли люди-общинники, мозг которых стал меньше по размеру, но в нем появились структуры, ответственные за речь, мышление и высокую координацию движений. То есть в данном случае можно говорить о формировании у представителей живой материи двух мощных полюсов адаптивности: «печеночного» адаптивного типа, более характерного для животных при наличии также мощного «мозгового» адаптивного типа характерного для троглодитов — неандертальцев. Такое сочетание открывало широкие возможности к развитию тормозных процессов в мозге и формированию более совершенных адаптивных механизмов, усиление которых лежало в основе формирования иерархии наиболее молодых структур коры серого вещества мозга, ответственных за становление и развитие ассоциативно-мыслительных процессов. Таким образом, появляется и все более совершенствуется механизм психологической защиты — адаптации, в основе которого противостояние суггестивному воздействию неандертальцев, дающее мощную энергетику биополя первобытной общины кроманьонцев. Судя по всему, ископаемые на сегодня формы людей достаточно долго сосуществовали с неандертальцами и сохраняли некоторые особенности их адаптивности, то есть были одновременно и магами и охотниками.
Происходило острое противостояние между теми и другими. Однако по мере развития человеческого общества (троглодиты-неандертальцы вымерли за счет утраты адаптивности) и перехода его в современную форму (человек разумный) это противостояние по генетическим и социально-энергетическим механизмам трансформируется в духовно-энергетическое поле, объединяющее социальные группы общества. В то же время формирование социально — энергетического поля создает определенную внутрипопуляционную иерархию с образованием различных социальных групп, формирующихся при распаде естества первичной социальной матрицы (крестьянской или кочевнической общины) народов, еще не ступивших в полной мере на путь процесса цивилизационного развития. А затем, по мере цивилизационного развития, противоборствующих социальных групп и классов.
У истоков человеческой мысли лежат остатки, следы ведической традиции. На сегодня ее классификация людей подразумевает их деление на 4 основных касты, сохранившихся в Индии. Согласно кастовой теории, человеческое общество имеет свою естественную иерархию. Определенные ступени этой иерархии могут занимать следующие общественные группы:
1) мыслители (брахманы), во многом определяющие самоидентификацию общества и направление его развития;
2) представители власти, силовых структур (кшатрии) определяющие субъектность, волю, во многом общественную энергию того или иного общества;
3) люди, обеспечивающие материальное благополучие общества — рабочие, инженеры, торговцы, земледельцы и т.д. — (вайшьи);
4) шудры — слуги всех трех высших групп.
Мы еще выделяем 5) группу — доисторический реликт — крестьянскую общину, след первобытной общины, которая в значительной степени сохранилась в русском дореволюционном крестьянстве, жившем, как известно, в техническом и культурном отношении в каменном веке, что и позволило ему сохранить отношения того времени в почти первозданном, естественном, изначальном виде. Это общество не имеет каких-либо иерархических структур, которые появляются лишь в процессе исторического развития тех или иных народов. В то же время сегодня совершенно неизвестно, какие свойства мозга позволяют формировать тот или иной тип общества – иерархию или общину.
Мы считаем, что экономика и производственные отношения имеют вторичный характер в развитии цивилизации. Базовый характер имеет тип развития и условия формирования центральной нервной системы, причем под условиями формирования мы понимаем не только внешние по отношению к организму факторы, но и такие внутренние факторы, как тип обмена веществ в организме, который в свою очередь зависит от общественных отношений. Отношения между людьми рождают определенную культуру как форму адаптации к окружающему миру. Важно определить какую культуру может породить тот или иной тип отношений, и содействовать ее развитию. Современная Россия совсем недавно, по историческим, меркам вышла из преобладания естества крестьянской общины, поэтому на данном историческом этапе мы не можем быть носителями подлинной буржуазной культуры, нам еще необходимо пройти фазу преобладания феодально – кшатрийской духовной структурализации. Такой этап развития был предсказан нам великим испанским философом Х. Ортегой-и-Гассетом («Восстание масс» 1937).
1. МЕХАНИЗМЫ АДАПТАЦИИ КАК ФАКТОРЫ НЕПРЕРЫВНОГО СОВЕРШЕНСТВОВАНИЯ ЖИВОЙ МАТЕРИИ.
1.1 МЕТАБОЛИЧЕСКАЯ «РЕКА» ФИЛОГЕНЕЗА
На заре появления живой материи при высокой температуре, давлении, радиации и других жестких условиях появились первичные органические молекулы: углеводы, жиры, аминокислоты. Конденсация этих веществ привела к появлению первичной протоплазмы с первичным метаболизмом, протекание которого целиком зависело от состояния окружающей среды, параметры которой и обеспечивали необходимую энергию протекания метаболических процессов в коацерватах (первичных функциональных единицах живой материи) а затем в первичных одноклеточных организмах (Опарин Л.И., 1958 г.).
Однако в дальнейшем условия на планете постепенно стали изменяться с уменьшением температуры, давления, радиации при увеличении количества углекислого газа, воды, водорода, серы и других веществ. В этих условиях у первичного органического материала стали все более и более проявляться отличия от неорганической материи, состоящие в способности модифицироваться и видоизменяться, сохраняя постоянство внутренней среды при изменении внешних условий.
Протекание метаболических реакций при снижении температуры, давления, радиации и других внешних мощных воздействий было возможно лишь при появлении белков-ферментов, резко понижающих энергетический порог этих реакций, т.е. энергия жестких внешних воздействий аккумулировалась в энергии полипептидной цепочки аминокислот, взаимодействующих друг с другом в виде полипептидных связей и составляющих полипептиды и белки. Ферменты и другие структурно-функциональные единицы живой материи включают в себя элементы неорганических веществ, то есть, неживой материи — железо, медь, кобальт, никель и др., входящих в активные центры белков — ферментов.
Моделирование условий, в которых согласно теории Опарина-Холдейна зарождалась жизнь на земле приводит к абиогенному синтезу 4-х аминокислот, входящих в состав белков всех организмов — это глицин, аланин, глутаминовая и аспарагиновая аминокислоты. Математический анализ современного генетического кода позволил выдвинуть предположение, что эти аминокислоты являются родоначальниками других аминокислот. Вступая во взаимодействие с соединениями серы, ароматическими веществами, претерпевая молекулярные перестройки, образуется все разнообразие известных на сегодня аминокислот. Аминокислоты выполняют выдающуюся роль в реализации метаболизма и функционирования органов и систем организма. Многих из аминокислот и их производные (ГАМК, ГОМК и тд.) являются адаптогенами, т.е. веществами, выполняющими защитные функции или принимающие участие в регуляции защитно-адаптивных систем организма. Что касается непосредственного участия, это антиоксидантный потенциал, защита воздействия от свободных радикалов, последствий воздействия радиации, в процессах обезвреживания токсичных веществ. Это энергетический и пластический ресурс организма. Они стабилизируют водно-электролитный баланс, клеточные мембраны и их ионные насосы (Кричевская А.А. 1983, Гончаренко Е.Н., Кудряшов Ю.Б. 1985, Xочачка П. 1988, Хазен А.М. 2000, Меерзон Ф.З 1984, 1999, 2001, Уайт А. 1981, Березов Т.Т. 1969.).
Неся на себе положительный и отрицательный заряд, а так же, обладая кислотными и основными свойствами, они поддерживают электролитный баланс, буферные системы, рН, а так же другие важные гомеостатические системы клеток и тканей. Необходимо отметить, что в состав белков многоклеточных организмов входят только L изомеры аминокислот, тогда как микробные клетки могут содержать D изомеры. С участием аминокислот (аргинина, метионина, глутамина и др.) происходит формирование систем дифференцировки и защиты организмов. ( Шлегель Г. 1986 )
По мере продвижения по филогенетической лестнице у организмов происходит упорядочение набора аминокислот и включение их в алгоритм реализации адаптивных механизмов. Так, у беспозвоночных животных фонд аминокислот чрезвычайно разнообразен и малоизучен. В тканях насекомых встречается огромное количество аргинина, пролина, глицина и глутаминовой кислоты, а так же амидов — глутамина и аспарагина, которые могут составлять более половины фонда свободным аминокислот. Целесообразность такого набора аминокислот у насекомых до сих пор остается не ясной. То же самое относится к таурину, глицину у морских моллюсков. ( Хочачка П., Сомеро Дж. 1977, 1988)
У высших животных содержание аминокислот приобретает тканевую и органную специфичность, строго постоянную для каждого вида. Кровь является связующим звеном между органами и системами в плане обеспечения и перераспределения аминокислот. Непрерывно протекающие процессы анаболизма и катаболизма белков определяют положительный или отрицательный азотистый баланс, характерный для процессов роста, адаптивных процессов при стрессовых воздействиях и патологических состояниях. В норме у взрослого организма наблюдается устойчивое равновесие аминокислот (Градусов Ю.Н. 1979).
Известно, что отдельные аминокислоты при определенных условиях сами собираются (спонтанно) в полипептидные цепочки, которые затем скручиваются во вторичные и третичные структуры — способные уже проявлять свойства белков (Уайт А. 1981).
Из данных литературы и наших собственных работ следует, что аминокислоты, помимо того, что они являются кирпичиками построения белков, биологически активных веществ в организме и других его важнейших структур, выполняют важнейшую функцию создания метаболического (обменного) фона, который определяет адаптивные возможности организма и их регуляцию. Т.е. соотношения аминокислот и их биологически активных производных в тканях организма являются своеобразной математикой живой материи, от изменений пропорций которой зависят защитно-адаптивные механизмы, регулируемые соотношением аминокислот (Павлов В.А. 2000, 2001, 2002).
Различные комбинации этих метаболитов могут создавать огромное количество вариантов (миллиарды) метаболических типов, создавая специфику обмена веществ у того или иного организма, а также адаптивные и метаболические особенности различных видов. В то же время количество нуклеотидов, из которых строятся нуклеиновые кислоты (ДНК и РНК), лежащие в основе хромосомного аппарата клетки и его воспроизводства, составляет всего 4 основных и два добавочных, из которых может образоваться 64 варианта кодонов, то есть основных элементов, которыми записывается генетический код. Учитывая приведенные обстоятельства и наши знания о воспроизводстве наследственной информации, как и филогенетической информации вообще, следует признать, что они еще очень недостаточны, для того, чтобы пытаться искусственным или полуискусственным путем без непредвиденных отрицательных последствий создавать новые организмы (клонирование, трансгенный перенос, попытки найти гены, ответственные за те или иные заболевания, и излечить эти заболевания путем отключения или стимуляции участков хромосом и т.д.). Появляются весьма аргументированные точки зрения, что генетика в современном ее состоянии вообще является лженаукой, и истинное ее развитие является делом будущего, при достижении человечеством более высоких научных и мировоззренческих горизонтов (П. Луизи 1993, Ф. Капра 2004).
Хорошо известен факт, что содержание свободных аминокислот в тканях обладает высоким постоянством и специфичностью, то есть для каждого вида животного, или даже отдельной особи, оно строго постоянно в его тканях (Уайт А. и др. 1981). Изменение количества аминокислот в тканях сопряжено со значительными перестройками защитно-адаптивных механизмов и выраженные отклонения наблюдаются при тяжелых патологических процессах (различных заболеваниях, травмах, старении и т.д.). Сопоставление содержания свободных аминокислот в тканях различных животных говорит о том, что у животных, находящихся на более ранних ступенях филогенетического развития общее содержание аминокислот в тканях больше, чем у животных, находящихся на более высоком уровне развития. В то же время качественный состав, разнообразие биологически активных веществ-производных аминокислот и ряда самих аминокислот, обладающих такой активностью, возрастает по мере филогенетического совершенствования организмов (метаболическая река филогенеза) (Хочачка Д.Ж., Самеро П. 1977, 1987, Кричевская А.А. 1981, 1982, Панин Л.Е. 1983, Западнюк В.И. 1983).
Особенно наглядно это видно по изменениям количества таурина и глютаминовой кислоты. Так, содержание таурина наиболее значительно в тканях организмов, стоящих на более древних, примитивных филогенетических уровнях. Затем по мере филогенетического совершенствования и появления высокоорганизованных защитно-адаптивных систем, количество таурина в тканях постепенно снижается, а глютаминовой и аспарагиновой аминокислот, триптофана, фенилаланина и их производных — увеличивается (Раевский К.С., Георгиев В.Л. 1986, Чирков А.Н. 1989, Кричевская А.А. 1981, 1982, Западнюк В.И. 1983).
И самое большое количество таких биологически активных веществ наблюдается в тканях приматов и человека, особенно в их головном мозге. Установлено, что стрессовые воздействия сопровождаются повышенным образованием метаболитов-адаптогенов в нейроэндокринной системе. В частности, в нейронах коры головного мозга образуются такие вещества как эндорфины, энкефалины, производные ряда аминокислот (глютаминовой, аспарагиновой, триптофана, фенилаланина, гистамина), являющиеся гормонами любви, счастья, ненависти, страха, воли и других проявлений духа, имеющего воплощение в организме, материи, обществе. (Судаков К.В. 1997, 2004, Хазен А.М. 2000)
Возможно, именно так и шел процесс адаптации первичной живой материи к изменяющимся условиям на планете, при использовании колоссального модификационного ресурса аминокислот. В этом плане следует отметить, что нуклеотиды — основные компоненты нуклеиновых кислот хранителей наследственной информации является по сути дела производными нескольких аминокислот: глицина, аспарагиновой кислоты, глутамина (Березов Т.Т.1969, Уайт А 1981).
А важнейшие регуляторы биосинтеза нуклеиновых кислот и пролиферативных процессов вообще — полиамины производные глутамата (орнитина, аргинина), метионина, лизина. Другие важнейшие регуляторы метаболизма: катехоламины, гормоны щитовидной железы (тироксин, тиронин), гистамин и серотонин, глутатион и карнозин (анзерин), а также многие другие биологически-активные вещества являются производными аминокислот (Кричевская А.А. 1983). Таким образом, важнейшими регуляторными метаболитами защитно — адаптивных систем организма являются либо сами аминокислоты либо их производные.
Поэтому вполне возможно, что свойства первичного метаболического субстрата во многом определялись особенностями аминокислот, входящих в него, а при изменении внешних факторов изменялись и эти особенности в сторону превращения аминокислот в регуляторные метаболиты. Изменение соотношения аминокислот-адаптогенов и их биологически активных производных, видимо лежало в основе совершенствования адаптивных механизмов.
Все это могло резко повышать приспосабливаемость органической материи к меняющимся условиям среды. И вершиной этого процесса стало появление из азотистых метаболитов нуклеиновых кислот и хромосомного аппарата. Все это предопределило формирование клеток и координированного воспроизводства филогенетической информации. Однако не следует переоценивать значение нуклеиновых кислот и хромосомного аппарата — это лишь верхняя ступень регуляторной пирамиды метаболического гомеостаза — средние и нижние ступени в ее памяти сохранились и заняты они белками (полипептидами), олигопептидами и аминокислотами. И вся эта информация воспроизводится в процессе полового и бесполого размножения, составляя метаболическую реку, в которой ключевое значение имеют взаимоотношения и особенности обмена важнейших аминокислот-адаптогенов.
Приводим строение 20 главных аминокислот необходимых для построения белков и биологически активных регуляторных метаболитов (рис 1. цит. по учебнику Березов Т.Т., Коровкин Б.Ф. 1990).
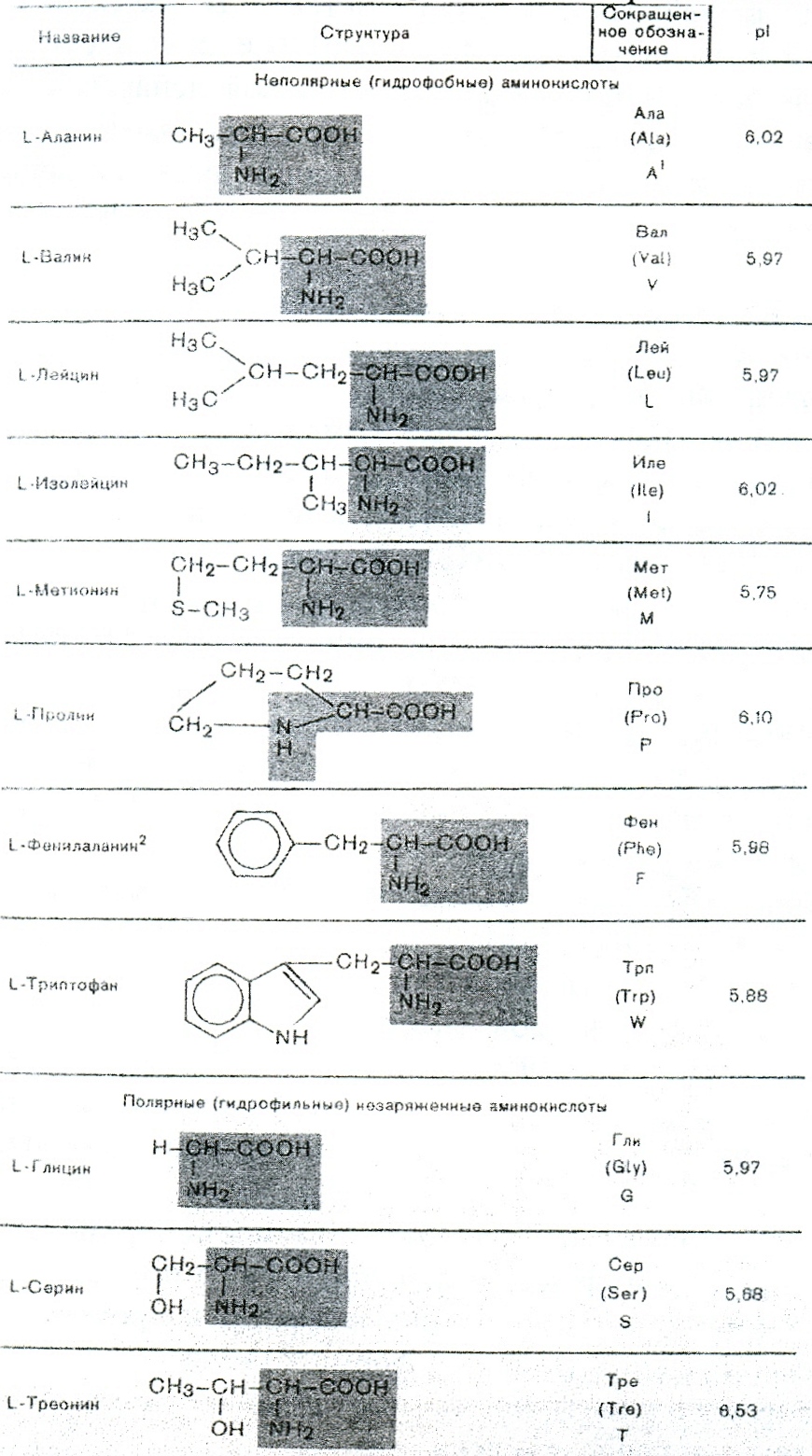

В настоящее время насчитывается более 200 различных производных основных аминокислот, что в самых различных комбинациях может нам дать колоссальное количество информации, из которой наиболее ценная, в той или иной стресс-ситуации, отбирается на уровне фенотипа и только затем фиксируется на уровне генотипа. Это можно подтвердить теми фактами, когда адаптивный ответ включается без структурного следа адаптации.
Имеются различные свидетельства того, что наследственная информация содержится и передается не только через гены, входящих в нуклеиновые кислоты. Наиболее известный пример: телегония. Друг Чарльза Дарвина лорд Мортон решил вывести суперпороду лошадей, скрестив английскую скаковую с жеребцом зебры. Но потомства не получил. Однако у родившихся затем у этой лошади жеребят от жеребца той же породы на туловище отчетливо были видны полосы как у зебры. Или другой пример: если породистую белую голубку скрестить с беспородным сизарем, то потом уже чистопородистого потомства у этой голубки уже не будет никогда, у голубят будут темные перья или какие-то другие признаки беспородных голубей. Иными словами, дети бывают похожи на любовников своей матери.
Можно привести еще один пример: если кошку скрещивать с ее собственным выросшим детенышем, то первые два помета у нее будут более менее нормальные котята, а в последующих пометах появляется все больше уродств, вплоть до того, что весь помет становиться нежизнеспособным. Если затем эту же кошку скрещивать с чужим котом, то в первых пометах также отмечаются нежизнеспособные уродливые котята и лишь затем, в четвертом — пятом помете котята становятся нормальными, но все равно количество уродств у котят такой кошки больше, чем у нормальной.
Все это говорит о том, что наследственная информация передается не только через хромосомы, а, возможно, через тот метаболический фон, который во многом может определяться спектром аминокислот — регуляторов-адаптогенов.
Аминокислоты в этом информационном потоке могут выполнять роль неспецифического базисного метаболического фона различных уровней филогенетической лестницы. Адаптивные механизмы, начиная с примитивных форм жизни, базируются на обмене аминокислот и их соотношениях, определяющих заряды и структурные элементы протоплазмы, получение энергии, стабилизации солевого обмена, рН и других важнейших параметров функции клетки ( Хочачка П..1988, Уайт А 1981, Лабори А 1972).
Наибольшее регуляторное значение имеют аминокислоты-адаптогены, обогащенные азотом (глутаминовая, глутамин, аргинин и ряд других) или имеющие в своем составе серу (метионин, цистеин, таурин и некоторые другие). Важнейшее регуляторное значение имеют производные аминокислот-олиго и полипептиды: полиамины, гаммаглутамилпептиды, имидазолпептиды и ряд других (Кричевская А.А. 1983, Хочачка П. 1988). Это значение является следствием того, что азот и сера имеют переменную валентность и могут подвергаться различной степени окисления, то есть менять свои химические свойства. Азот имеет на своих энергетических уровнях 7 электронов, причем на последнем — 4 электрона; сера имеет на своих энергетических уровнях 16 электронов и на последнем 6 электронов. Из чего следует, что сера имеет больший диапазон окислительно-восстановительных превращений и поэтому неслучайно, что в процессе эволюции природа выбрала именно серу, как важнейший инструмент в регуляции механизмов обмена веществ в живой природе. Так, азот может окисляться от 0 валентности (N2 азот воздуха) до +5, например, в составе аммиака. Сера окисляется от 0 валентности (ископаемая сера) до +6 в серистом ангидриде или даже до -2 в сероводороде. Отмеченный диапазон окислений, определяет образование различных метаболитов, которые участвуют в регуляции различных метаболических процессов. Так, например, восстановленная сера (SH) является ключевым метаболитом для поддержания нормального функционирования многих ферментов организма, а таурин (содержащий SО3) является мощным антиоксидантом и стабилизатором мембран; остаток серной кислоты (SО4). Окисленная сера входит во многие структурные компоненты клеток, например, она необходимая для образования соединительной ткани — гепарин, хондроитин-4-сульфат, хондроитин-6-сульфат, то есть, кислых гликозамингликанов. Имеются сведения, что соотношения кислых и нейтральных гликозаминогликанов определяют различия метаболизма и функциональных состояний важнейших адаптивных систем организма (кроветворная ткань, лимфоидная ткань) (Юшков Б.Г.,1992).
Большое значение имеет количество и состояние окисленности железа и серы в активных центрах белков ферментов (Fe-S) ( Хьюз М. 1983 ). В свою очередь, повышение в тканях метаболитов, обогащенных азотом (глутамин, полиамины, нуклеиновые кислоты) отчетливо стимулируют пролиферативные процессы. Поэтому соотношение содержания серы в тканях и степени ее окисленности с одной стороны и азота с другой стороны являются важнейшим регуляторным механизмом в направленности метаболических адаптивных особенностей организма — N и S тип . Но сера и азот в тканях представлены в соответствующих серо и азотсодержащих аминокитслотах. Таким образом, из имеющихся данных следует, что в адаптивных механизмах преобладают N или S аминокислоты.
Совершенствование форм жизни сопряжено с изменениями азотистого обмена и ресурсов аминокислот — переход от аммониотелии к урикотелии и уреотелии (Хочачка П. 1988).
Исходя из концепции пифагорейцев, окружающая нас природа выражена в определенных соотношениях, достигающих определенного уровня гармонии, в наибольшей степени это проявляется в живых организмах, существующих вопреки неблагоприятным условиям окружающей среды, благодаря высокой степени организации живой материи. Наивысшей степени совершенства достигают аэробные организмы, прежде всего человек, использующие для своей жизнедеятельности окисление поступающих с пищей веществ кислородом. Однако это же обстоятельство определяет повреждающее его воздействие в условиях стрессовых нагрузок, особенно превышающих адаптивные ресурсы организма.
Кислород и его активные радикалы могут повреждать все ткани организма, особенно его многофункциональных систем: нейроэндокринной, дыхательной, сердечно-сосудистой, выделительной и других, что и лежит в основе патогенетических нарушений при многих патологических состояниях, в том числе и при старении. По сути, весь организм можно рассматривать как высокоорганизованную антиоксидантную систему, предназначенную для сохранения информации, содержащейся в половых клетках. Исходя их этих представлений защитно-адаптивныe возможности организма, его здоровье и продолжительность жизни во многом зависят от состояния его антиоксидантных систем, обеспечивающихся, в свою очередь, необходимыми ресурсами N и S аминокислот.
У млекопитающих все регуляторные метаболиты либо сами являются аминокислотами, алигопептидами, полипептидами, либо производными аминокислот. Все это создает некий метаболический порог-стресса, выход, за который означает развитие дистрессового состояния или патологию. Накапливается все больше фактов о том, что возвращение к физиологическому регуляторному уровню (структурный след адаптации) можно осуществить введением в организм, который подвергается какому-либо запредельному воздействию, ряда аминокислот-адаптогенов (глутамата, различные полиптиды и смеси аминокислот, полиаргинина, цистеина, метионина, олигопептидов — тафтсин, тимоген).
Соматические клетки могут делиться 50-70 раз, в то время как половые клетки могут делиться бесконечно, то есть обладают практически бессмертием, их существование поддерживается их окружением или всем организмом, в которых эти половые клетки образуются. Так, организмы выполняют защитно-адаптационные функции по поддержанию метаболического потока сконцентрированного в половых клетках. При слиянии мужских и женских половых клеток происходит метаболический взрыв, сопровождающийся расширением метаболической информации. Расширение это доходит до стадии половозрелого состояния организма, а затем начинает постепенно уменьшаться вплоть до полной утраты его адаптивных возможностей. На смену исчерпавшему свои защитно-адаптивные возможности приходит другой организм.
С этих позиций процесс зачатия новой жизни в материнском организме, это не есть только слияние Х и У хромосомы, а встреча двух информационных потоков, сохраняющих в своей памяти все этапы филогенетического развития того или иного вида и все этапы предшествующего филогенеза живой материи.
Исходя из этих представлений рождение, развитие, зрелость и старость любого организма можно представить как восприятие от своих родителей информационного потока филогенетической информации, зафиксированного на определенном уровне в виде спектра и определенного фонда аминокислот и их соотношения, что в стратегическом плане зафиксировано в виде генетической информации в генах. Стресс-воздействия могут существенно менять оперативный уровень метаболизма или фонотип того или иного индивидуума, но на генетическом уровне эта информация откладывается лишь в случае адекватности воздействий метаболическому уровню той или иной ступени филогенетической лестницы.
На ранних стадиях онтогенеза плод пользуется метаболическим фондом матери, на который накладывается вводимый с семенем метаболический фонд отца ( Цанев Р.Г., Марков Г.Г. 1964 ).
По мере созревания плода меняются пропорции метаболического фонда, что предопределяет основные параметры роста и развития организма. По мере взросления осуществляется некий баланс в системе N/S. Однако если на ранних этапах развития преобладает S тип по мере старения организма, возрастает доля N типа. Метаболическая память организма отличается от генетической своей высокой мобильностью и при неблагоприятных воздействиях, превышающих стрессовый порог, возможен откат на более древние метаболические системы (например, при массивной кровопотере или травматическом шоке начинает преобладать аммониотелический тип азотистого обмена, характерный для беспозвоночных). Это позволяет пережить повреждающее воздействие и затем выйти на физиологический уровень. Введение в организм подопытных животных аминокислот-адаптогенов, например, глутаминовой кислоты, значительно ускорит этот процесс, что препятствует развитию необратимых патологических процессов (Волков М.С и др. 1975. Павлов В.А. и др. 2000.).
В этом плане, необходимо указать на то, что эксперименты по клонированию животных и методы генного инженеринга не могут быть успешными и безопасными без учета этих обстоятельств, ибо полноценная филогенетическая информация несется не только с генами но и со всей той средой, которая окружает половые клетки. Так, сперма содержит большое количество полиаминов и богатых азотом аминокислот: аргинана, лизина, таурина, в то время как готовая к оплодотворению слизистая матки содержит большое количество серосодержащих аминокислот, серы и глутатиона и богатых SH группами белков и метаболитов (Алов И.А., Брауде А.И., Аспиз М.Е. «Основы функциональной морфологии клетки» 1966. ).
В ходе оплодотворения и дальнейшего развития плода меняется метаболический фон как вокруг плода, так и в женском организме вообще. Изменяются пропорции аминокислот с повышенным количеством азота (N — глутамин, аргинин, полиамины и некоторые другие) и серы (S — метионин, цистеин, таурин и ряд других), регуляторных метаболитов N и S типов. После рождения плода изменение этих пропорций продолжается вплоть до достижения половозрелого состояния. Этот период у разных классов и видов является тем периодом, когда организм в результате различных внешних воздействий способен максимально к ним приспосабливаться, т.е. включаться в филитический поток информации, вносить в него свою лепту и передавать далее по наследству.
С возрастом, испытывая на протяжении жизни повреждающие (стрессовые) воздействия, меняется метаболический ресурс организма. Эти изменения у аэробных организмов, использующих в окислительно-восстановительных процессах (ОВП) кислород, связаны, прежде всего, с тем, что снижаются антиоксидантные ресурсы и антирадикальные ресурсы вообще. Т.е. защитные механизмы, направленные на предохранение клеток, и тканей (прежде всего мембранных структур) от повреждения кислорода и его активных форм (АФК), возникающих в ходе его использования в качестве окислителя в обменных процессах (Арчаков И.И. 1975, Афонасьев И.Б. 1988).
Таким образом, заложенная в метаболическом фонде программа подразумевает в процессе жизнедеятельности организма постепенное «стирание» этого фонда о стресс — воздействия или точнее повреждающее воздействие 02 и особенно образующиеся в процессе метаболизма его активные формы в меняющихся условиях среды. Изменение метаболического фонда ведет к дисбалансу в системе.
Процессы старения у человека касаются, прежде всего, нервной ткани, особенно коры головного мозга и соединительной ткани, т.е. структур, ответственных за обеспечение метаболических процессов организма аминокислотами, входящими в семейство глутамата и серосодержащими аминокислотами-адаптогенами. При этом происходит прогрессирующее снижение SH групп в регуляторных белках — ферментах и других важнейших адаптивных системах организма, ответственных за их функциональное состояние, с последующими нарушениями в азотистом, жировом, углеводном обмене и развитием тех или иных патологических состояний старения (Анисимов В.А. 1981, Джиоев Ф.К., Плис Б.Г. 1984).
Диспропорции в ресурсах регуляторных N- и S-аминокислот помимо периода старения характерны и для периода созревания организма. Однако, если в период старения преобладают и прогрессируют N-системы со склонностью к воспалительно-дистрофическим пролиферативным (опухолевым) процессам, то в детском возрасте преобладает S-тип с гиперконтролем над пролиферативными процессами и относительной редкостью дистрофически-опухолевых процессов в этот период (Раевский К.С. 1986, Нефедов Л.И. 1999).
Избыток серосодержащих адаптогенов защищает головной мозг ребенка от гипоксии и родовой травмы. Однако преобладание S-типа делает организм ребенка менее устойчивым к большинству инфекций и токсических воздействий. Такое преобладание, очевидно, обусловлено тем, что голова и мозг ребенка имеют значительно больше размеры по отношению к другим частям тел, чем у взрослого человека. При этом интенсивность метаболических процессов: окислительно-восстановительных и образование серосодержащих аминокислот и глутамата наиболее интенсивны в раннем детском возрасте в головном мозге ребенка с постепенным снижением этих параметров по мере роста и взросления организма. А в процессе старения идут обратные процессы с уменьшением объема мозга, интенсивности в нем ОВП, образования глутамата и серосодержащих метаболитов. Болезни, приводящие к снижению ОВП мозга, сопровождается снижением образования в нем глутамата и серосодержащих аминокислот и угнетением адаптивных возможностей организма (болезнь Альцгеймера, Паркинсона, Дауна и др.).
1.2 АМИНОКИСЛОТЫ АДАПТОГЕНЫ В МЕХАНИЗМАХ РЕГУЛЯЦИИ ЖИЗНЕДЕЯТЕЛЬНОСТИ КЛЕТКИ
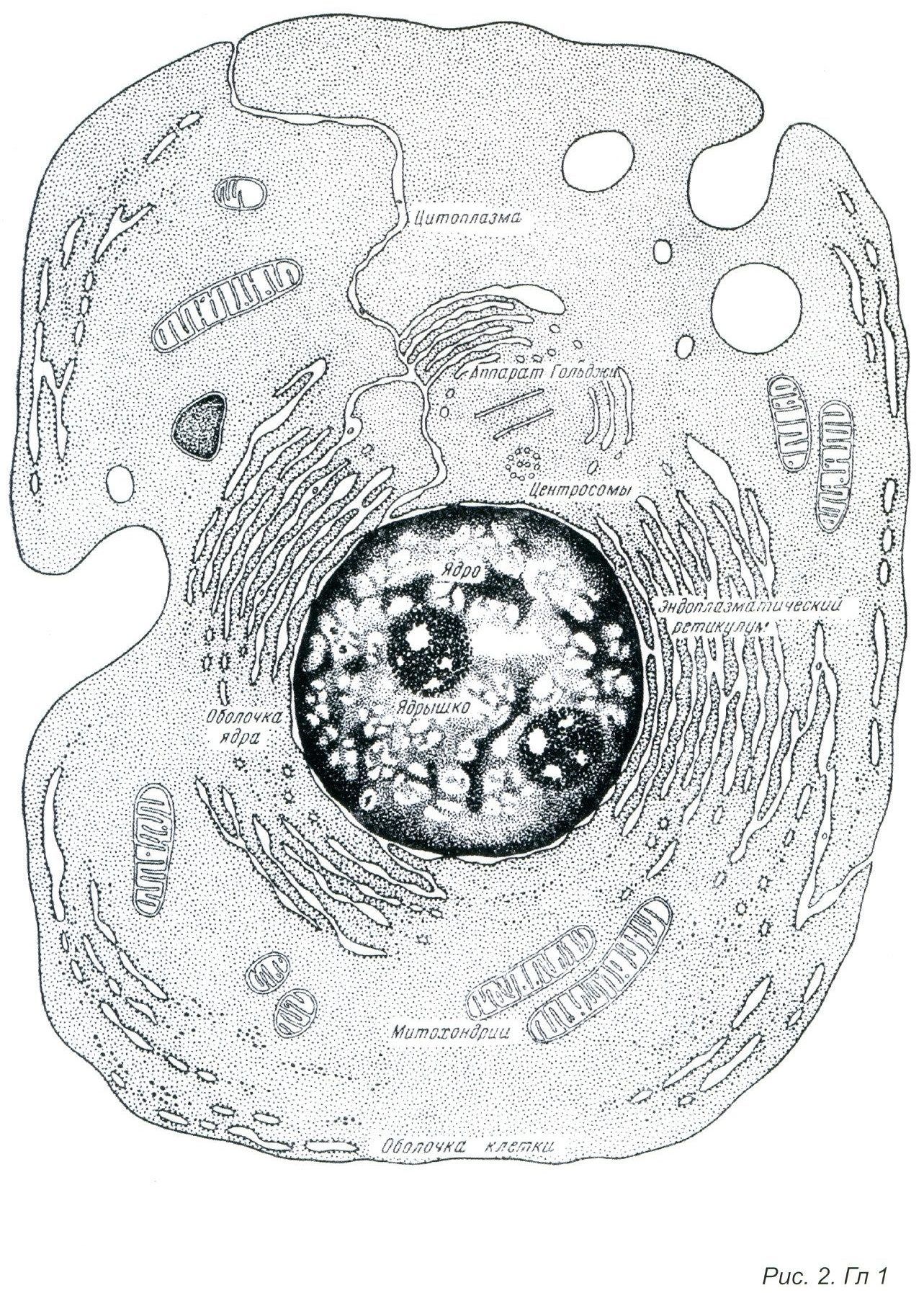
Начать обобщение наших знаний об адаптивных механизмах живой материи необходимо с элементарной ячейки живой материи-клетки. На приведенном рисунке можно видеть ряд важнейших структур клетки. Важнейшими органеллами, во многом определяющими адаптивные перестройки клетки, являются митохондрии, в которых протекает окисление поступающих в клетку питательных веществ (углеводы и жиры). В результате чего образуется энергия АТФ, выделяется углекислый газ и вода. Окислительно-восстановительные процессы (ОВП) в митохондриях протекают при интенсивном использовании кислорода. (Рис. 2)
На сегодня установлено, что для процессов жизнедеятельности организмов, а, соответственно, и для защитно-адаптивных механизмов необходима энергия, которая накапливается в клетках в виде макроэргического соединения АТФ (аденозинтрифосфат), или в виде ГТФ (гуаназинтрифосфат). В дальнейшем АТФ используется для самых разнообразных структурно-функциональных перестроек и динамической работы и других функциональных процессов клеток и тканей. ГТФ используется в процессах биосинтеза, то есть в построении новых биологических структур.
АТФ можно рассматривать как первый медиатор-гормон, появившийся на заре зарождения живых организмов (Хазен А.М. 2000). Имея в своем составе адениловый нуклеотид и остатки фосфорной кислоты АТФ обладает многочисленными регуляторно-защитными функциями начиная от способности аккумулировать и отдавать энергию и выполняет различные регуляторные функции как на уровне мембран, так и на уровне цитозоля клетки. Соотношение АТФ-АДФ-АМФ влияет на многие стороны жизнедеятельности клетки, прежде всего ее энергетические функции. Важное регуляторное значение имеют производные АТФ и ГТФ циклические нуклеотиды — ЦАМФ и ЦГМФ, регулирующие пластические и функциональные процессы в клетке. Для образования энергии в клетках и тканях необходимо поступление в них достаточного количества кислорода. Следует отметить, что большинство организмов используют в своих ОВП в качестве окислителя кислород (аэробные организмы). Но есть организмы, живущие в бескислородной среде и использующие в своих ОВП другие окислители, например серу, азот некоторые кислоты. Но наиболее эффективное образование энергии и возможность активного функционирования отмечается именно у аэробных организмов использующих кислород. То есть в филогенетическом плане это более совершенные и адаптивные организмы (Шлегель Г. Худолей В.В. 1993, 1997).
Процессы образования энергии являются неспецифическими, т.е. необходимы для всех организмов в их адаптивных перестройках и ничем у аэробных организмов не отличаются. Они протекают абсолютно одинаково у человека, животных, простейших и бактерий, использующих кислород.
Окислительно-восстановительные процессы в клетках происходят в нескольких формах:
1. Окисление поступающих с пищей углеводов, жиров и белков (аминокислот, которые могут превращаться в углеводы в процессе глюконеогенеза) происходит в митохондриях клеток при участии группы ферментов объединенных в так называемый лимонно-кислый цикл или цикл Кребса. В регуляции функции митохондрий ключевое значение имеют системы сопрягающие азотистый и энергетический обмен. Процесс переаминирования и дезаминирования аминокислот и включения их в энергетические процессы или напротив, образования аминокислот, происходят с участием трансаминаз аланина и аспарагиновой кислоты, а так же фермента — глутаматдегидрогеназы. Кроме того, имеются шунтовые механизмы с участием аспарагиновой кислоты и аланина, сопрягающие процессы метаболизма в митохондриях и цитозоле клеток, Т.е. ОВП тесно связано с обменом аминокислот и находится под контролем соотношения аминокислот — адаптогенов. В условиях повышенных нагрузок на организм важное энергетическое значение имеют аминокислоты, образовавшиеся вследствие усиления протеолитических процессов. Из которых, преимущественно в печени, образуется глюкоза — процессы глюконеогенеза. Строго говоря, стресс реакция на метаболическом уровне во многом сводится к усилению протеолитических процессов за счет выброса глюкокортикоидов и обеспечения организма необходимой энергией за счет реакций глюконеогенеза (Кендыш И.Н., Голиков П.И. 1988 ).
Важнейшим веществом, которое образуется при таком окислении в митохондриях является НАДН2 (никотинамиддинуклеотид восстановленный). В такой промежуточной форме накапливается энергия окисления перечисленных выше питательных веществ. Затем, от НАДН2 отщепляется водород и электроны, с помощью дыхательной цепи (группы окислительно-восстановительных ферментов, находящихся в митохондриях), то есть там же, где протекает цикл Кребса. В результате окислительно-восстановительных процессов в дыхательной цепи образуется универсальное энергетическое соединение АТФ (аденозинтрифосфат), обеспечивающее большинство энергоемких процессов в тканях организма, углекислый газ и вода, которые выделяются из организма легкими и почками.
Необходимо отметить, что НАД, так же как и АТФ, является важнейшим защитно-регуляторным метаболитом, обеспечивающим регуляцию как энергетических, так и пластических процессов за счет способности к окислению и восстановлению. Соотношение восстановленных и окисленных его форм во многом определяет механизмы регуляции. Присоединение фосфата дает другой важный защитно-регуляторный метаболит — НАДФ. Таким путем спектр защитно-регуляторных возможностей этих метаболитов существенно возрастает. Необходимо отметить, что как пуриновые, так и пиримидиновые нуклеотиды, входящие в состав АТФ и НАДФ, являются производными нескольких аминокислот: аспарагиновой, глицина, глутамина и наличие этих аминокислот в клетках во многом влияет на образование перечисленных нуклеотидов и их отношение (Хазин А.М. 2000).
2. Другой группой окислительно-восстановительных процессов в клетке является так называемый пентозный цикл, протекающий в цитоплазме клетки. В этом цикле окисляются углеводы (глюкоза) с образованием НАДФН2 (никотинамиддинуклеотидфосфат восстановленный). Этот процесс не требует кислорода, а образующийся НАДФН2 необходим для поддержания процессов биосинтеза вновь образующихся структур клетки, особенно при их делении и росте. Также большое значение пентозный цикл имеет в обеспечении обезвреживания (детоксикации) попадающих в организм чужеродных веществ (ксенобиотиков) или структур тканей и клеток, подвергающихся повреждению, которые необходимо выводить из организма (эубиотики). В регуляции многих сторон жизнедеятельности клетки большое значение имеют регуляторные метаболиты, имеющие в своем составе SH группы, прежде всего это глутатион. Поддержание этих групп в восстановленном состоянии обеспечивается системами образования НАДФН2 (пентозный цикл, работа маликфермента) и образование аскорбиновой кислоты в соединительнотканных структурах организма. Функции этих систем тесно взаимосвязаны и, как показано в наших исследованиях, регулируются метаболитами обмена глутаминовой кислоты: оксипролин, аргинин и ряд других, т.е. так же зависимы от обмена аминокислот.
3. Еще одной группой ОВП являются процессы обезвреживания ксено- и эубиотиков ферментами, располагающимися на гладком эндоплазматическом ретикулуме, то есть системе оксигеназ смешанных функций (ОСФ) находящихся на мембранах, как бы сложенных в цитоплазме клетки, то есть ОВП с использованием кислорода протекают и в цитоплазме клетки. Это необходимо для инициации детоксикационных процессов при окислении чужеродных веществ (ксенобиотиков), попадающих в клетку, или обезвреживании токсичных продуктов, образующихся в тканях в ходе обмена веществ (эубиотики). В дальнейшем, после окисления, эти вещества соединяются с обезвреживающими метаболитами, такими, как глутатион, некоторые аминокислоты, и ряд других веществ с дальнейшим выведением из клеток, тканей и организма.
Имеются еще некоторые группы окислительно-восстановительных процессов, например, прямое окисление липидов в пероксисомах.
Для нормального протекания окислительно-восстановительных процессов необходимы следующие условия: как уже отмечалось, обеспечение в достаточном количестве тканей организма кислородом с одной стороны, с другой необходима адекватная данной нагрузке антиоксидантная защита. В случае, если эти процессы сбалансированы, то адаптивные системы организма способны выводить его на уровень, устойчивый к тому или иному повреждающему воздействию. Если необходимого соответствия в этих процессах нет, то возникает либо гипоксическое состояние (нехватка кислорода), либо состояние пониженной антиоксидантной защиты. В том и в другом случае снижается образование носителей энергии в клетке АТФ и ГТФ, нарушается ее функция, а затем функции тканей и системы в целом. Длительное воздействие повреждающего фактора с понижением АОЗ приводит к развитию «окислительного стресса» с повреждением мембран и других структур клеток и тканей, с нарушением их функций. В случае продолжения повреждающего воздействия, к их гибели (Арчаков А.А. 1975, Афанасьев И.Б. 1988, Лукьянова Л.Д. 1982).
Кислород — это мощный окислитель, который является достаточно агрессивным веществом, способным повреждать мембраны и другие структуры клеток, контактирующих с ним. Еще более агрессивными являются промежуточные продукты, образующиеся из кислорода в ходе ОВП. К таким продуктам относят супероксидный анион, синглетный кислород, перекись водорода и ряд других веществ, способных атаковать окружающие мембраны и структуры клеток. В норме процессам избыточного образования активных форм кислорода (АФК), токсичных для клетки, препятствует антиоксидантная защита (АОЗ) клеток и тканей. Эта система включает в себя несколько белков-ферментов (супероксиддисмутаза, каталаза, пероксидаза, глютатионредуктаза и глютатионпероксидаза), а также различные вещества, способные препятствовать повреждающему воздействию кислорода и его АФК. К таким веществам относят витамины А, Е, К и С, многие гормоны и биологически активные вещества. А также ряд других простых и сложных соединений. Однако необходимо отметить, что многие из перечисленных метаболитов и сложных соединений при интенсивном и часто повторяющемся стрессовом воздействии на ткани и клетки утрачивают антиоксидантные возможности, а зачастую, и сами становятся прооксидантами и агрессивными веществами, способными повреждать мембранные структуры. Например, гормоны (катехоламины), биологически активные амины (серотонин, гистамин), витамины (витамин С) и ряд других веществ. И только аминокислоты, особенно серосодержащие (цистеин, таурин), а также содержащие повышенное количество азота (глутамин, аргинин, лизин) до конца выполняют свою антиоксидантную функцию. Еще более мощный антиоксидантный защитный эффект оказывают некоторые производные перечисленных аминокислот: глутатион, полиамины, а также ряд других веществ, содержащих в своем составе несколько отмеченных выше аминокислот (олигопептиды). (Кричевская А.А. 1983).
Другой важнейшей структурой клетки — является ядро, в значительной мере определяющее специфический, присущий только этому организму ответ на повреждающее воздействие. В ядре сосредоточена генетическая информация в виде спирали ДНК, находящаяся в хромосомах. Эта информация может воспроизводиться за счет информационных РНК, которые передают заложенную в генах информацию на рибосомы клетки (расположенных на мембранах эндоплазматического ретикулума — шероховатый эндоплазматический ретикулум). На рибосомах происходит биосинтез белков- ферментов, необходимых для протекания любых обменных процессов в организме. Рибосомы — состоят из рибосомальной РНК и белков, они являются своеобразными фабриками по биосинтезу вновь образующихся белков. В этом процессе используются 20 основных аминокислот, которые как кирпичики складываются в первичную структуру белка. Затем силы электрофизического и химического взаимодействия скручивают эту последовательность в спираль — вторичную структуру белка, которая затем складывается под воздействием этих же сил, а также физико-химических факторов окружающей среды в третичную структуру — собственно сам белок. Далее такие субъединицы могут соединяться в функционально-активные белковые объединения, например, некоторые ферменты, состоящие из нескольких субъединиц. В последовательности аминокислот, а также во всех последующих структурах белковой молекулы заложена колоссальная информация, которая реализуется в ходе различных обменно-адаптивных превращений (Березов Т.Т. 1990, Уайт А. и др. 1981).
Считается, что в регуляции воспроизводства наследственной хромосомной информации большое значение имеют гормоны (соматотропный, глюкокортикоиды, инсулин). Кроме того, имеется обширная, но противоречивая литература о влиянии на генетический аппарат количества SH групп, АФК, свободных радикалов. Однако, какой либо определенной и убедительно обоснованной концепции о регуляции генетического аппарата продуктами метаболизма до сих пор не существует. Большинство работ, посвященных функции генетического аппарата и биосинтеза белков в клетках выполнены на Е.соli. У многоклеточных организмов эти процессы идут значительно сложнее, и многие стороны их еще мало изучены. Поэтому возможности эффективно модулировать эти процессы пока нет. (Хазин А.М. 2000, Ф. Капра, 2004).
Исходя из данных литературы и результатов наших исследований, аминокислоты-адаптогены, влияющие на биосинтез таких важных регуляторов метаболизма как пуриновые и пиримидиновые нуклеотиды, глутатион и другие биологически активные олиго и полипептиды, например, тафтсин, холицистокинин, гистоны, полиамины, находясь в определенном соотношении, соответственно, влияют на функцию генетического аппарата клетки. Иными словами, они могут сопрягать неспецифическую и специфическую фазы стресс-реакции с последующим образованием необходимых белков — ферментов, выводящих организм из фазы тревоги в фазу резистентности.
Так, белки выполняют регуляторную, рецепторную, защитно-опорную, а также другие функции, обеспечивающие специфический ответ клетки на внешние воздействия и изменение обменных процессов под этими воздействиями.
Наибольшее значение для выполнения структурно-опорной функции имеют белки соединительной, костной ткани, сосудов, нервных волокон. Специфические функционально-морфологические особенности организма во многом определяются иммунной системой, в которой образуются белки — антитела и антигены. Иммунная система выполняет защитную функцию по отношению к чужеродным элементам (например, возбудитель инфекционных заболеваний) устраняя их из организма, а также элиминируя собственные измененные и поврежденные клетки и ткани.
Белки играют значительную роль в регуляции обменных процессов организма, так они встраиваются в мембраны клеток, определяя их функциональное значение. Работа митохондрий, ядра, рибосом, обезвреживающих механизмов на гладком эндоплазматическом ретикулуме выполняются соответствующими белковыми структурами.
Как ферменты, белки обеспечивают протекание всех обменных процессов в организме. В последовательности аминокислот и их сборке во вторичную, третичную, четвертичную структуру, заложена колоссальная информация, которая накапливалась миллиарды лет в процессах эволюции живой материи. Достаточно сказать о том, что протекание реакций обмена веществ без наличия белков-ферментов потребовало бы температуры в сотни и тысячи градусов, высокого давления и других, несовместимых с физиологическим уровнем клетки условий. Поэтому можно сказать, что в цепочке аминокислот, из которых построены белки, заключена информация и энергия тех процессов, которые определяли развитие филогенеза с момента появления живой материи на планете. Однако, помимо того, что аминокислоты выстраиваются в определенную последовательность в белках, или более коротких молекулах — полипептидах или олигопептидах, они также находятся в свободном состоянии во всех клетках и тканях организма и выполняют важнейшую регуляторную функцию. Как мы уже отмечали, соотношение этих свободных аминокислот строго постоянное и меняется лишь под воздействием интенсивных повреждающих факторов или значительных адаптивных перестройках организма. (Кричевская А.А. 1982, Западнюк В.И. 1983, Уайт А. и др. 1981).
Все это позволяет заключить, что соотношение аминокислот как в молекулах белков или полипептидов, так и в свободном состоянии является (в терминологии пифагорейцев) сакральной геометрией живой материи или тем математическим языком, который регулирует эту материю, через который окружающая среда взаимодействует с организмами или популяциями этих организмов. Таким образом, аминокислоты, создавая структуры белка (первичную, вторичную, третичную, четвертичную), а также создавая различные соотношения в клетках и тканях в живой материи во многом определяют ее энергетически-адаптивный потенциал и, изменяясь в результате внешних воздействий, эти соотношения регулируют защитно-адаптивные механизмы.
1.3 ВАЖНЕЙШИЕ ЗАЩИТНО-АДАПТИВНЫЕ СИСТЕМЫ ОРГАНИЗМА
Для контроля за адекватным протеканием адаптивных перестроек к изменениям окружающей среды в организме имеются неспецифические защитно-адаптивные системы.
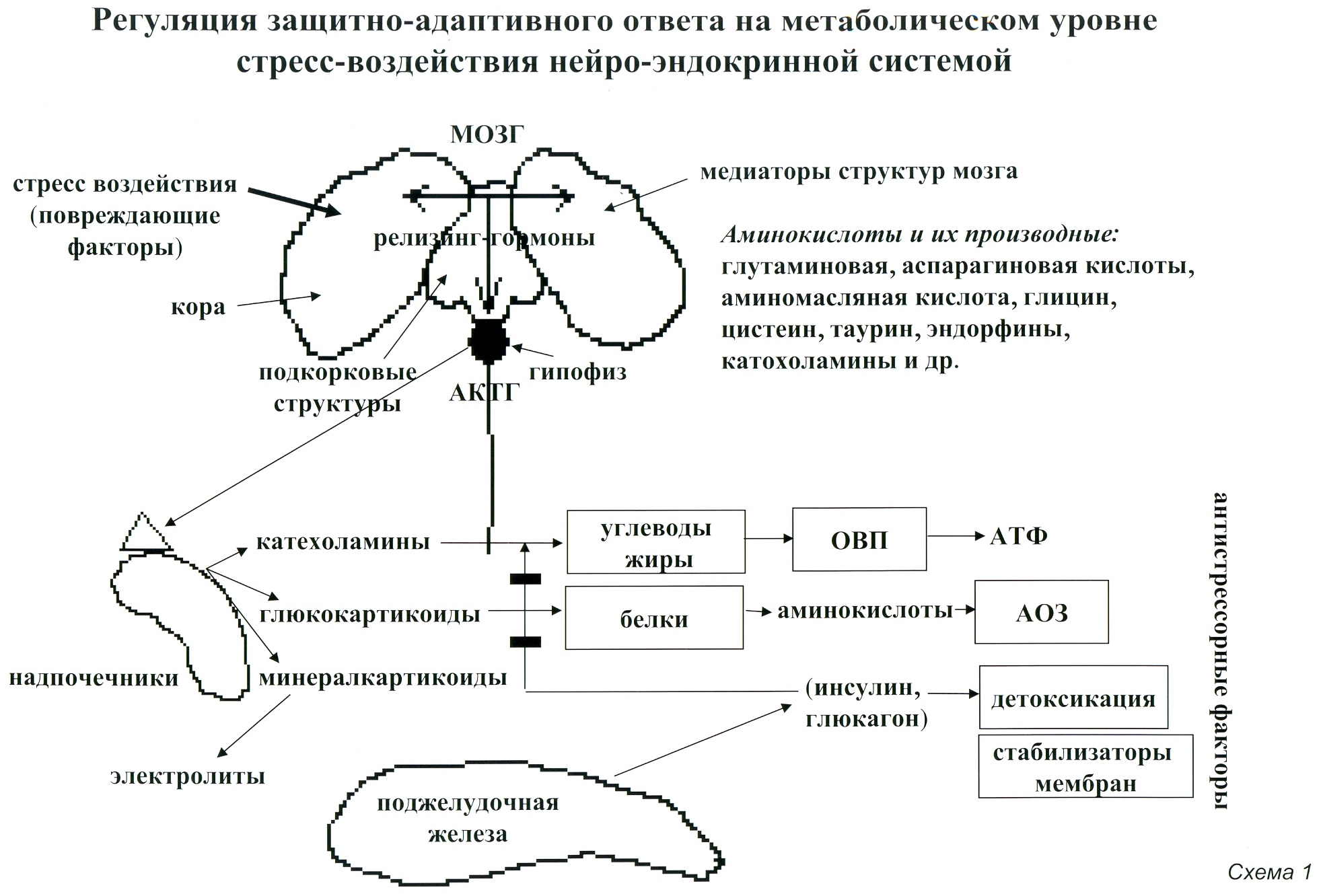
1. Нейроэндокринная. Эта система включает в себя мозг, в котором выделяют кору и подкорковые структуры, а также продолговатый и спинной мозг и железы внутренней секреции (гипофиз, щитовидная железа, надпочечники, поджелудочная железа, половые железы).
В нейронах в условиях стресс-воздействия образуется большое количество РНК, биологически активных белков, полипептидов, что во многом определяет формирование процессов памяти (Меерзон Ф.З. 1996, 2001). Значительное усиление метаболических процессов сопряжено с избыточной энергообеспеченностью, которая необходима для реализации адаптивных процессов (Меерзон Ф.З. 1998, Хазин А.М 2000.). При этом в головном мозге в больших количествах образуются такие метаболиты как аммиак, мочевина, глутаминовая и аспарагиновая аминокислоты. Все эти вещества являются мощными регуляторами защитно-адаптивных механизмов (Козлов Н.Б.1971, Кричевская А.А. 1983, Меерзон Ф.З.1997).
Важнейшее значение в регуляции нейроэндокринной системы, особенно ее высших отделов (кора головного мозга и подкорковые структуры), имеют некоторые аминокислоты-адаптогены. К возбуждающим аминокислотам — медиаторам относят глутаминовую и аспаргиновую кислоты и ряд их производных, а также триптофан и фенилаланин, терозин и их производные (катехоламины (КХА), гормоны щитовидной железы — тироксин и другие). К тормозным аминокислотам — медиаторам относят гамма-аминомасляную кислоту — ГАМК, таурин, глицин. (Раевский К.С., Георгиев В. Л. 1986, Волков М.С. 1975, Whight C.Е. 1986)
Нейроэндокринная система имеет главенствующее значение в адаптивных перестройках организма, как к воздействиям внешней среды, так и внутренним изменениям организма. Внешние воздействия воспринимаются и обрабатываются корой головного мозга, затем информация передается на подкорковые структуры (гипоталамус, гипофиз). В гипоталамусе вырабатываются биологически активные вещества — релизинг-факторы, которые поступают в гипофиз. Далее в гипофизе вырабатываются тропные гормоны, например адренокортикотропный гормон, который имеет важнейшее значение в реализации механизмов стресса. Он стимулирует выработку надпочечниками важнейших гормонов стресса: катехоламинов (КХА) и глюкокортикоидов (ГК). КХА значительно усиливают расщепление углеводов из гликогена («депо» углеводов в клетках), а также мобилизуют жиры из их «депо» в органах и подкожно-жировой клетчатке. В результате в крови резко повышается уровень углеводов (глюкоза) и жиров (жирные кислоты), которые поступают в клетки и интенсивно окисляются в их энергетических центрах-митохондриях. Здесь образуется энергия в виде АТФ, необходимой для стимуляции адаптивно-защитных механизмов и повышения устойчивости к повреждающему стресс-воздействию (Голиков П.П. 1988, Голиков С.И. 1986, Кендыш Л.А. 1986).
Глюкокортикоиды способствуют расщеплению белков в ряде тканей организма, подвергающихся разрушающему воздействию стрессового фактора: лимфоидная ткань, соединительная, мышечная, жировая ткань, некоторые ткани желудочно-кишечного тракта, и некоторые другие. Смысл такого разрушения состоит в резком увеличении аминокислот-адаптогенов для стимуляции защитных механизмов, противостоящих процессам повреждения и стимулирующих восстановительные биосинтетические процессы (реконструктивный протеолиз), что в целом повышает устойчивость организма к повреждающему фактору (Покровский А.А. 1976). (Схема 1)
2. Сердечно-сосудистая система. Эта система включает в себя сердце, выполняющее механическую функцию и сосуды различного диаметра. Эта система выполняет функцию транспорта кислорода (в эритроцитах), от легких к тканям, где поступающий кислород включается в ОВП. А также транспорта других необходимых для обеспечения жизнедеятельности веществ (углеводов, жиров, аминокислот, витаминов, электролитов и других).
3. Дыхательная система: верхние дыхательные пути (бронхи) и легкие, которые обеспечивают проникновение кислорода из вдыхаемого воздуха в кровеносные сосуды, а затем в эритроциты.
Имеются сведения, что в регуляции этих систем на метаболическом уровне большое значение имеет образование оксида азота и соответствующие ему ферменты. Образование оксида азота и его регуляторное значение на защитно-адаптивные системы организма тесно связанные с обменом аммиака, глутаминовой кислоты, аргинина и мочевины. В тканях имеются различные NO синтазы (Е-эндотелиальные, Н-нейрональные, I-индуцибельная), которые определяют наработку оксида азота в эндотелии, нейронах и фагоцитирующих клетках. С определенным уровнем оксида азота связаны функциональные возможности этих систем (Проскуряков А.Н. 2000, Каминская Г.О. 1999.
4. Желудочно-кишечный тракт обеспечивает поступление пищевых веществ и их расщепление до метаболитов, способных всасываться через кишечную стенку в кровь и транспортироваться в виде углеводов, жиров, аминокислот к тканям в качестве необходимого топлива для протекания ОВП и процессов биосинтеза.
В желудке и кишечнике вырабатываются биологически активные пептиды такие как холецистокинин, серотонин, гистамин и ряд других оказывающих влияние на эмоционально волевую сферу, процессы памяти, сна. Наработка этих веществ влияет на процессы интеграции между адаптивными система организма. Печень является своеобразной биохимической лабораторией организма, образующей большое количество биологически активных адаптогенов.
5. Кроветворная — сосредоточена в трубчатых и плоских костях в виде костного мозга и необходима для постоянного образования клеток крови: лейкоцитов, эритроцитов, тромбоцитов, выполняющих различные защитно-адаптивные функции в организме.
6. Иммунная система включает в себя железы регуляции иммунитета (вилочковая железа — тимус), а также периферические ткани иммунной системы (селезенка, лимфатические узлы различных органов, тканей и сосудов).
Имеются данные, что в регуляции этих систем так же имеют большое значение аминокислоты, принимающие участие в реализации регуляторных механизмов (цистеин, таурин, глутамигновая и аспарагиновая аминокислоты, метионин, аргинин, пролин) ( Белокрылов Г.А 1988, Павлов В.А., Сабадаш Е.В. 2001, 2002).
7. Антиоксидантная система — система, отвечающая за предотвращение повреждающего воздействия кислорода, его промежуточных метаболитов на важнейшие структуры клетки, прежде всего мембраны (на чем мы останавливались выше).
8. Выделительная система это система обезвреживания вредных веществ поступающих в организм извне (ксенобиотики), а также поврежденных и измененных структур и молекул собственно организма — эубиотиков. Начинается такое обезвреживание в метаболических системах клетки.
Первая и ключевая реакция обезвреживания в метаболических системах связывание аммиака, образующегося в высокофункциональных тканях, прежде всего в нейронах головного мозга. Обезвреживание происходит при связывании аммикака глутаминовой кислотой с образованием глутамина и выведением его за пределы клетки.
На мембранах гладкого эндоплазматического ретикулума (ГПР) ксенобиотики или образующиеся в организме вещества подлежащие обезвреживанию, подвергаются окислению в системе оксигеназ смешанных функций (ОСФ), а затем связыванию полученных метаболитов с некоторыми аминокислотами (таурином, глицином, глютамином) а также некоторыми специально синтезируемыми веществами (глутатионом, уридиндифосфоглюкуроновой кислотой (УДФГК) и фосфоаденозинфосфосульфатом (ФАФС) и выводятся из клетки через аппарат Гольджи, представляющий из себя конечные отделы эндоплазматического ретикулума. Эти процессы наиболее интенсивно протекают в клетках печени, легких и менее интенсивно в коже, клетках крови и клетках других тканей. Затем обезвреживаемые вещества в виде соединений с перечисленными выше метаболитами выделяются в кровь и далее с мочой, потом и другими выделениями выделяются из организма.
Отмеченные неспецифические зашитно-адаптивные механизмы, рассматриваемые нами на уровне отдельных организмов универсальны, но живая материя входит в единую систему-биоценоз, который также можно рассматривать, как своеобразный организм с присущими ему защитно-адаптивными системами и механизмами развития. Эти особенности реализуются на видовом, межвидовом, популяционном и других уровнях биоценоза. Существует различные биологические цепи ,определяющие как качество жизни, так и самое существование того или иного вида. Так, в пищевой цепи, травоядные зависят от наличия растительной пищи, хищники зависят от количества травоядных. Погибающие организмы как травоядных, так и хищников служат пищей для простейших и бактерий, преобразующих их останки в азотистые удобрения, и увеличивают массу растительной пищи и т.д. Другим примером биологической цепи может быть трансгенный перенос между различными видами животных и человеческих популяций, осуществляемый микроорганизмами (простейшие, бактерии, вирусы). Микромир имеет очень высокий потенциал адаптивных перестроек и очень чутко реагирует на изменение окружающей среды (как глобальные космические воздействия, так и отрицательные последствия деятельности человека), поэтому трансгенный перенос осуществляется микроорганизмами, переносящими метаболически-генетическую информацию как между отдельными особями, так и между различными видами, что является важнейшим адаптивным механизмом биоценоза (Шлегель Г. 1987, Л. Маргулис 1988).
Существуют разнообразные формы симбиотических (взаимовыгодных) отношений между организмами. Так, некоторые виды птиц уничтожают паразитов на теле и в ротовой полости китов, крокодилов, бегемотов. Последние, в свою очередь, служат защитой для этих птиц.
Давно известна неблаговидная роль таких паразитов, как гельминты. Однако в последние годы установлено, что и они выполняют важную функцию в биоценозе. Показано, что заражение ими может приводить к излечению, например, такого заболевания как неспецифический язвенный колит. Обычные методы лечения этого заболевания не всегда эффективны и иногда прходится прибегать к хирургическому удалению всего толстого кишечника.
Глисты, как и другие паразиты, оказывают воздействие на НЭС, кроветворную, иммунную, защитно-адаптивную системы, способствуя их перестройке. Но во многом их взаимодействия нам не понятны.
Еще большее значение для регуляции защитно-адаптивных систем макроорганизмов имеет микрофлора: бактерии, простейшие, спирохеты, грибы, вирусы и другие ее представители. Микроорганизмы распространены повсеместно, во всех слоях биосферы и контактируют со всеми населяющими ее организмами, активно воздействуя на их защитно-адаптивные системы. Так хорошо известен факт, что у человека на поверхностях, контактирующих с внешней средой, находится большее количество микроорганизмов, чем его собственных клеток, составляющих различные органы. И все эти микроорганизмы выполняют какую-то свою симбиотическую функцию. Например, микрофлора кишечника, вырабатывает основное количество необходимых для организма витаминов группы В — В1, В6, В12 и ряд других важных компонентов для нормального протекания обменных процессов (Шлегель Г. 1986, Уайт А. 1981).
В случае гибели нормальной микрофлоры, например при некоторых заболеваниях желудочно-кишечного тракта, развивается дисбактериоз с чрезмерным размножением патогенных бактерий, грибов. Нарушается обеспечение организма витаминами, в организм попадают токсичные вещества, может развиться тяжелое нарушение обмена веществ, вплоть до гибели организма, что чаще отмечается у детей. Введение необходимых микроорганизмов (бифидумбактерии, лактобактерии, кишечная палочка) препятствуют развитию и прогрессированию заболевания.
В наших исследованиях (Павлов В.А. и соавт. 2000, 2001) было установлено, что микобактерии туберкулеза существенно снижают мутагенное воздействие ароматических ксенобиотиков на экспериментальных животных.
Согласно симбиотической концепции (Шлегель Г. 1986, Готтшалк Х, 1980), важнейшие энергетические структуры клетки-митохондрии являются потомками бактерий, которые взаимодействовали с первичными многоклеточными оранизмами в анаэробный период биосферы. Это повышало адаптивные возможности тех и других. В дальнейшем, произошло их слияние в единую систему. Имеются точки зрения, что помимо митохондрий тканей, микротрубочки нейронов мозга так же имеют бактериальное происхождение. Кроме того, метаболический аппарат оксигеназ смешанных функций в наибольшей степени выраженный в печени, так же, возможно ,имеет бактериальное происхождение. Однако, если бактерии,послужившие родоначальниками митохондрий современных организмов, обладали преимущественно ферментными системами, способными к метаболическим превращениям с участием кислорода, то бактерии или грибобактерии, давшие начало оксигеназам смешанных функций, имели мощные ферментные системы, включавшие в себя цитохром Р 450. Последние в большей степени были способны интенсивно метаболизировать ксенобиотики тиап ПАУ.
Определенным подтверждением такой концепции можно считать отсутствие цитохрома Р-450 и системы ОСФ почти на всех ступенях эмбрионального развития большинства животных и человека. Необходимо отметить, что каждый индивидуум проходит все ступени филогенеза в своем онтогенезе и только после рождения и по мере взросления организма количество цитохрома Р-450 резко увеличивается в тканях (печени, легких, кожи).
Примечательно, что образование стероидных гормонов в надпочечниках и половых железах, для образования которых также необходим цитохром Р-450, также начинается после рождения плода, Т.е. после резкого усиления аэробных процессов в организме. Все это наводит на мысль о том, что образование стероидных гормонов — своеобразный эндогенный механизм регуляции, который в какой-то мере дополняет или конкурирует с регуляторными воздействиями экзогенных биорегуляторов-ПАУ. Регуляция эта может быть связана с тем, что стероиды, так же как и ПАУ, которыми они по сути дела и являются, так как в их основе лежит молекула циклопентанпергидрофенатрена, обезвреживаются или подвергаюся метаболическим превращениям по тем же путям, что и ПАУ. Или иными словами ПАУ можно рассматривать как экзогормоны, а образующиеся в организме метаболиты сходные с ними по строению как эндогормоны или ксенобиотики и эубиотики.
Эти вещества легко проникают в липиды организма, изменяя их свойства, т.е. свойства мембран (Голиков Н. С. 1886, Нечипоренко Н.П. 1981). Подвергаясь обезвреживанию в системе ОСФ, эти вещества могут давать метаболиты, ковалентно связывающиеся с нуклеиновыми кислотами и другими нуклеофильными и активными центрами биологических молекул. Это обуславливает мутагеные и канцерогенные воздействия ПАУ ,т.е., вызывающие мутации — изменение генетической информации и развитие патологических процессов вплоть до опухоли, нарушение ОВП клеток и ряд других нарушений метаболизма, связанных с образованием АФК и агрессивных метаболитов ПА У и ПОЛ.
Согласно представлениям, сформулированным учеными в последние годы (Л. Маргулис, П.Л. Луизи) микромир играет исключительную роль в интеграции биоценоза. Установлено, что микроорганизмы могут трансформироваться друг в друга, утрачивая свою прежнюю специфику и приобретая новые свойства. Ежедневно микромир обменивает до 15% своей генетической информации. При этом он осуществляет трансгенный перенос этой информации, проникая в различные макроорганизмы и взаимодействуя с их генетическими структурами. И мы можем добавить, что микромир взаимодействует и с метаболическими защитно-адаптивными системами макроорганизмов, таким путем интегрируя их на уровне видов, популяций, биоценоза в единые защитно-адаптивные системы. (Схема 2)

Согласно концепции Чижевского А.Л. («Эпидемические катастрофы и периодическая деятельность солнца» М. 1930), микромир имеет большое значение для развития видов животных и популяций человеческого общества и соответственно цивилизации. Так эпидемии и пандемии таких опасных заболеваний как чума, оспа, холера, туберкулез имеют определенную цикличность во времени и влияют на процессы цивилизационного развития.
1.4 РОЛЬ АМИНОКИСЛОТ — АДАПТОГЕНОВ В МЕХАНИЗМАХ РЕАЛИЗАЦИИ СТРЕСС — РЕАКЦИИ
В 30-40 годах прошлого столетия Г. Селье была разработана теория стресса. Она описывает универсальное свойство живой материи при воздействии на нее различных повреждающих факторов. Но несмотря на большие успехи в области биохимии, физиологии, молекулярной генетики и других направлениях, достигнутых с того времени, что якобы, позволяет даже клонировать не только животных, но и человека и разрабатывать другие методы воздействия на генотип, во многом эти попытки сродни средневековой алхимии.
Однако если алхимик подвергал риску только собственную голову, в данном случае опасность подстерегает человечество в целом, так как отдаленные результаты этих эспериментов практически предсказать нельзя ( Ф.Капра. 2004). В механизмах адаптации и реуляции адаптивных систем остается много неясного.
Во многом остается непонятно, как осуществляется координация между важнейшими гомеостатическими системами организма и генетическим аппаратом его клеток, не известны медиаторы, которые модулируют функцию генетического аппарата. Во всяком случае, в наших руках пока нет средств, позволявших бы активно вмешиваться в механизмы стрессового воздействия без отрицательных последствий для организма (П. Хочачка, Дж. Смеро. 1988). Согласно той же концепции Г.Селье, стрессовое воздействие имеет две формы: эустресс и дистресс. Эу — хороший и Дис — плохой. А также можно выделить гипостресс и гиперстресс.
Г. Селье выделяет три фазы стресса:
1. Фаза тревоги — когда происходит возбуждение НЭС. Интенсивно образуется большое количество биологически активных веществ — стрессфакторов, обуславливающих расщепление белковых структур и разрушение ряда тканей организма для обеспечения необходимого метаболического материала для процессов приспособления организма к повреждающему воздействию.
2. Фаза резистентности — повышенная устойчивость к повреждающему воздействию.
3. Фаза истощения — в случае повторных воздействий повреждающих факторов.
После прекращения действия чрезвычайного раздражителя в организме восстанавливается типичный для него базальный уровень стресс – гормонов: катехоламинов, глюкокортикоидов, инсулина, а также метаболический фон.
Однако, несмотря на то, что теория стресса подробно разработана и имеет многочисленных сторонников, по мере углубления знаний о механизмах повреждающего воздействия различных факторов на организм и о различных типах реагирования на эти воздействия, появляется все больше противоречий в трактовке механизмов адаптивного ответа на стресс воздействие. Это может быть связано как филогенентически обусловленными межвидовыми отличиями, так и вызванными различными патологическими состояниями нарушениями адаптивных механизмов. Более того, внутри каждого вида найдены существенные отличия в механизмах реагировании.
Такие известные физиологи, занимавшиеся проблемой адаптации как Л.А. Орбели, П.Д. Горизонтов, В.П. Казначеев, Ф.З. Меерзон, Л.Е. Панин и многие другие, не отрицая понятия стресс, предлагают свои схемы ответа организма на повреждающее воздействие. В результате возникает пестрая картина, которая не позволяет выработать единую теорию и алгоритм подходов для объяснения понятий стресс и адаптация и их взаимодействия.
Однако иногда спор доходит до отрицания самого понятия стресса, так У. Седерберг считает этот термин ничего не выражающим. Другие авторы (Ю.М. Губачев, К.В. Судаков) трактуют стресс только в контексте эмоций (П.В. Симонов, Л.Леви) или предлагают разделить стресс на эмоциональный, физический и смешанные варианты.
Другой важной проблемой является наличие специфических и неспецифических реакций при стрессе и их взаимоотоношения. Еще сам Г.Селье подчеркивал, что основные проявления стресс реакций не специфические, однако некоторые авторы подвергают сомнению неспецифический характер, так как каждый стимул имеет свой характерный ответ. Все это существенно затрудняет как понимание, так и коррекцию последствий перенесенных воздействий.
В работах отчественных исследователей (И.П. Павлов, Ф. З. Меерзон, Л.Е. Панин) обоснованно выделяются как неспецифические так и специфические факторы стресса.
Причем первые — удел фазы тревоги, когда влючаются реакции нейроэндокринной системы, мобилизация жиров, углеводов, аминокислот, катаболические процессы, что сопровождается образованием эндогенных стимуляторов, резко повышающих процессы энергетики и запускающих каскад метаболических процессов, направленных на повышение резистентности организма и выход его на адаптивный уровень. В исследованиях Ф.З. Меерзона показано, что неспецифические процессы в определенной степени формируют специфичный ответ, который во многом заключается в формировании памяти на уровне генетического аппарата клеток. Наибольшей емкостью этой памяти обладают клетки нейроэндокринной системы, прежде всего нейронов. В которых резко возрастает количество функционально биологически активных ДНК, РНК, олиго и полипептидов и аминокислот — нейромедиаторов. Высказывается мысль о том, что образующиеся в тканях мозга метаболиты — адаптогены непосредственно могут включаться в метаболические процессы на уровне поврежденных тканей (глутаминовая кислота, ГОМК, полипептиды и др. (Меерзон Ф.З. 1996, 1999, Хазин А.М 2000). Таким путем нервная система может оказывать регуляторное воздействие не только путем генерации нервного импульса, но и непосредственно за счет выхода метаболитов в кровеносное русло. В тоже время надо признать, что факторы, связывающие неспецифическое и специфическое звено стресса не найдены, что является слабым звеном теории стресса и нет достоверных научно обоснованных подходов для воздействия на реализацию стресс реакции в нужном нам направлении.
Биологический смысл стресса состоит в том, что он оставляет структурный след адаптации. Происходят изменения на генетическом уровне, усиливается биосинтез белков — ферментов, ответственных за перестройку метаболических систем, лежащих в основе функционирования важнейших адаптивных систем: нейроэндокринной, дыхательной, сердечно-сосудистой, выделительной, детоксикационной, антиоксидантной.
Согласно полученных Паниным Л.Е. и соавторами данных ключевое значение в развитии фазы резистентности при стрессе имеют особенности функции поджелудочной железы и печени. Что касается поджелудочной железы, то это выработка инсулина — важнейшего антистрессового гормона и поддержание его необходимых концентраций на определенном уровне, достаточном для противодействия гормонам стресса. В печени образуются липопротеиды высокой, низкой и очень низкой плотности, которые как показано автором, осуществляют регуляцию функции митохондрий в условиях стрессового воздействия и одновременно доставляют основные субстраты (липиды) к этим энергетическим центрам. Развитие фазы резистентности как раз во многом связано с переключением наработки энергии в тканях за счет преимущественного использования липидов, в процессах прямого окисления их в пероксисомах клеток. Тогда как в фазе тревоги преимущественно используются углеводы в процессах гликолиза и цикла Кребса.
Большое значение в особенностях протекания стресс — реакции имеет чувствительность организма к глюкокортикоидам, стимулирующим протеолитические процессы в условиях стрессового воздействия. Условно выделяют глюкокортикоид положительные и глюкокортикоид — отрицательные типы, что зависит от наличия или отсутствия в тканях- мишенях глюкокортикоид — рецепторных белково — полипептидных комплексов, подвергающихся в процессе стрессового воздействия протеолизу с образованием аминокислот- адаптогенов (Голиков П.П., 1986). К глюкокортикоид — положительным относится большинство млекопитающих, к глюкокортикоид- отрицаельным — морские свинки, приматы и человек. В печени образуется большое количество белков, которые присутствуют в крови (альбумины, белки острой фазы воспаления и другие полипептндные структуры) подвергающихся катаболизму в условиях стрессового воздействия. Однако у разных организмов, даже одного вида., а тем более у представителей различных видов, этот процесс имеет существенное различие. Таким образом, из имеющихся литературных данных можно сделать предположение, что существует различные адаптивные типы — с преобладанием первой фазы стресса (возбуждения) и соответственно гормонов этой фазы и второй фазы- (резистентности) с преобладанием гормонов поджелудочной железы и метаболитов, синтезируемых в печени.
Эти данные в определенной степени были подтверждены в исследованиях Панина Л.Е., Казначеева В.П, Маянского Д.Н. которые нашли, что как животные, так и люди подразделяются на «тревожных» и «спокойных» особей.
Все это предопределяет разный гомеостатический уровень адаптивных возможностей организма — повышающий или снижающий устойчивость к повреждающим воздействиям.
Важнейшей неспецифической основой изменений метаболизма при стрессе является перестройка окислительно-восстановительных процессов с усилением энергетического обеспечения адаптивных систем в организме. Стрессовое воздействие приводит к нарушению работы основных энергетических центров клеток-митохондрий. За счет повреждения их мембран и ферментных комплексов. Считается, что любое стрессовое воздействие в фазу тревоги сопряжено с генерацией в тканях организма большого количества активных форм кислорода (АФК). Это предопределяется повышенным образованием в хромофинных клетках надпочечников и других отделов НЭС — катехоламинов (КХА), нарушающих нормальное течение ОВП в высокоаэрируемых тканях (легкие, сердце, мозг, кости и др.). Образующиеся при стрессе в больших количествах катехоламины и глюкокортикоиды обезвреживаются в системе оксигеназ смешанных функций (ОСФ) в наибольшем количестве имеющихся в клетках печени, легких, кожи, ряда клеток крови. В процессе обезвреживания образуются промежуточные метаболиты этих веществ, АФК, которые могут повреждать мембранные структуры клеток. В то же время отмеченные процессы способствуют включению компенсаторных механизмов клеток, тканей и систем адаптации. Прежде всего, в тканях усиливается гликолиз, происходит мобилизация и расщепление жира из имеющихся ресурсов организма. Энергетический обмен переключается на использование жира — как более эффективное в условиях повышенной нагрузки и неблагоприятных воздействий на организм. Однако главенствующее значение в условиях повреждающих нагрузок имеет способность организма усиливать свою антиоксидантную защиту (АОЗ) (Панин Л.Е. 1983).
Необходимо еще раз подчеркнуть, что молекулярные механизмы стресса изучены недостаточно.
В монографии Панина Л.Е. «Биохимические механизмы стресса» 1983 мы находим следующие представления о механизмах стресса.
Известно, что в функционировании НЭС в роли медиаторов выступают КХА, ацетилхолин, серотонин, глутаминовая кислота (и ее производные), глицин, таурин.
На уровне ЦНС вырабатываются пептидные гормоны — релизинг — факторы, на уровне периферических желез НЭС — гормоны коркового и мозгового слоя надпочечников, В поджелудочной железе — инсулин и глюкагон. Кроме того, в различных тканях и органах вырабатываются тканевые биологически активные вещества — тканевые гормоны: циклические нуклеотиды, простагландины и лейкотриены, а также ряд других метаболитов. На уровне иммунной системы вырабатываются гормоны тимуса, цитокины. Известно, что все эти вещества реализуют свой эффект через рецепторные механизмы. Однако появляется все больше сообщений о том, что их эффект может быть связан с непосредственным включением компонентов этих биологически активных веществ в метаболические процессы.
В последние годы становится все более понятно, что образующиеся в организме свободные радикалы кислорода, азота, а также радикалы обезвреживаемых в тканях веществ экзогенного происхождения (ксенобиотики) и эндогенных метаболитов (эубиотики, например, катехоламины, глюкокортикоиды, производные холестерина, триптофана и ряда других веществ, а также продуктов разрушения мембран клеток, различных белков, липогликопротеидов и т.д.) могут также играть медиаторную функцию — стимуляции, угнетения и сопряжения различных систем организма при повреждающих воздействиях. ( Каминская Г.О 2004, Проскуряков А.Н. 2004, Сахно О.И. 2000 ). Именно эти промежуточные продукты метаболизма при стрессовых воздействиях могут выполнять роль того загадочного по Г. Селье вещества, которое реализует эффекты стрессового воздействия .
Кроме того, уже много лет, начиная с работ Богомольца, Ужанского известно, что в механизмах аварийного регулирования при гиперстрессе или дистрессе большое значение имеют эндогенные метаболиты — адаптогены — продукты катаболизма, Т.е. продукты разрушения тканей и клеточных структур, например, клеток крови (эритродиерез). Они обладают мощным стимулирующим воздействием на защитно-адаптационые системы организма: кроветворную, иммунную, антиоксидантную и другие. Так, мощным стимулирующим, восстанавливающим действием обладают продукты распада клеток при травме, воспалении, гипоксии. Это прежде всего полипептиды, олигопептиды и некоторые аминокислоты: глутаминовая, глутамин, аргинин, пролин, лейцин, валин и ряд других .
Все эти вещества можно объединить как эндогенные адаптогены организма. Однако, несмотря на то, что учение о стрессе существует уже более пятидесяти лет и многочисленные исследования посвященные этой проблеме, до сих пор так и не вырисовывается какой-то определенной картины или картин, позволявших бы количественно и качественно охарактеризовать метаболические сдвиги и главные регуляторные перестройки, позволявшие бы эффективно регулировать или вовсе заблокировать нежелательное стрессовое действие или, напротив, создать адекватные модели стрессовых воздействий.
Основная проблема, на наш взгляд, состоит в том, что не известны были те промежуточные фазы регуляции, которые сопрягают неспецифическое звено стресса (ОВП-ПОЛ-АОЗ) и его специфический компонент — реакции иммунного ответа, структурной перестройки на уровне генетического аппарата с последующим образованием на метаболически-генетическом уровне структурного следа адаптации, позволяющего нарабатывать необходимые белки — ферменты и биологически — активные метаболиты для реализации адекватной повреждающему воздействию стресс — реакции.
Мы считаем, что на эту роль могут претендовать аминокислоты и некоторые их производные. Стресс-реакция протекает по-разному у представителей различных ступеней физиологической лестницы. У человека большое место занимают стрессы, вызванные особенностями жизни и общения между людьми (психоэмоциальные стрессы, воздействие неблагоприятных факторов среды из антропогенных источников — загрязнение биосферы ксенобиотиками, шум, вибрация, заразные заболевания и т.д.), к чему организм человека имеет потенциальные механизмы защиты, начиная от метаболических систем обезвреживания вредных веществ и кончая интеллектом, призванным вырабатывать гармоничные формы существования человека и остальной биосферы.
У животных, стоящих на более низких, чем человек, ступенях филогенетической лестницы стресс-реакция на внешние воздействия менее разнообразна по формам защитных механизмов, участвующих в адаптивных перестройках.
Так, стоящие близко к нам млекопитающие не подвержены таким психоэмоциональным воздействиям как человек, ибо эти воздействия не являются для них адекватными в их жизни.
Как отмечается в монографии Панина Л.Е (там же), у пресмыкающихся земноводных, рыб стресс-реакция по своим механизмам отличается от таковой у млекопитающих.
На определенном этапе филогенеза, разделение первоначально единой нервной системы на вегетативную и соматическую предопределило возникновение трофического аппарата и органов движения.
Нервная система эволюционно связана с эндокринной. Родоначальниками медиаторов нейроэндокринной системы считаются нейрогормоны, такие как катехаламины, эфиры холина, которые обнаружены у бактерий, грибов простейших, растений.
У беспозвоночных уже имеется 2 типа нервной передачи: адренэргический и холинэргический, а также определенную роль в них имеет серотонин.
У низших позвоночных хромофинная ткань еще не сконцентрирована в симпатических нейтронах и образующиеся в ней адренэргические вещества осуществляют регуляцию гуморальным путем. Т.е., через выделение гормонов в кровь.
У высших позвоночных симпатико-адреналовая система заключает в себе два типа клеток-нейроны симпатической нервной системы — (образованиее норадреналина) и клетки хромофинной ткани (образующие адреналин). Важнейшие медиаторы cтpecc-реакции-стероидиые гормоны у рыб и амфибий не разделены на минерал- и глюкокортикоиды. Синтез альдостерна стал возможен только после выхода позвоночных на сушу.
Другим важным гормоном стресс-реакции является инсулин, который как бы противостоит глюкортикоидам и катехоламином в реализации стресса на уровне органов и тканей. Считают, что предшественники В-клеток поджелудочной железы имеются уже у беспозвоночных. У кишечнополостных их обнаружить не удается. У круглоротых и низших позвоночных инсулярный аппарат не сконцентрирован в одном месте и разбросан по печени и кишечнику. У костистых рыб В-клетки концентрируются уже в отдельном органе (брокмановское тельце). Окончательное формирование поджелудочной железы завершается у земноводных и пресмыкающихся.
Нейроэндокринные механизмы, наряду с аллостерическими (активизация или угнетение ферментов, обеспечивающих обменные процессы избытком или недостатком образующегося в этих процессах вещества) составляют фундамент регуляции всех функций организма, предопределяемых метаболическим фоном и создающий метаболический порог адаптивных возможностей организма, представляемых состоянием обмена веществ. Однако тонкая регуляция метаболизма имеет еще более устойчивую основу — своеобразный нулевой цикл, представляемый метаболическим фоном и создающий метаболический порог адаптивных возможностей организма. В зависимости от этого фона происходит реализация регуляторных эффектов гормонов и других регуляторов стресс-реакции.
Этот самый метаболический фон имеет решающее значение при выходе организма из пределов стресс-реакции в исходное состояние или даже повышение резистентности организма к повреждающему фактору и степени обратимости дистресса.
Т.е. можно констатировать, что в процессе филогенеза усложнение адаптивно — функциональных возможностей организма сопряжено с расширением набора биологически активных метаболитов (аминокислот и их производных) В механизмах регуляции адаптивных перестроек — «расширяющяяся метаболическая река филогенеза».
Практически все метаболиты живых клеток, обогащенные азотом: глутамин, аспарагин, аргинин, мочевина, полиамины, нуклеотиды обладают защитными по отношению к кислороду и АФК свойствами (Кричевская А.А 1983). Эти же вещества стимулируют пролиферацию клеток и рост тканей, Т.е., значительное усиление пластических процессов. Это есть своеобразная антиоксидантная защита. Возможно, что, огромная масса динозавров именно этим и объясняется. Как мы уже указывали, 60-100 млн. лет назад концентрация кислорода в атмосфере Земли была в 2-3 раза выше, чем сегодня, на чем мы более подробно остановимся ниже.
Однако повышение 02 в атмосфере за счет пышного и буйного роста растительности сотни миллионов лет назад вследствие интенсивного фотосинтеза приводило помимо появления древнейших животных-гигантов к пожарам в древних лесах, при горении которых образовывалось большое количество ксенобиотиков типа полициклических ароматических углеводородов (ПАУ). Воздействие большого количества 02 и ПАУ могло оказывать мощное мутагенное воздействие на древнейших животных, чем, вероятно, и объясняется их поразительное многообразие. Однако со временем содержание 02 в атмосфере стало снижаться, а ксенобиотиков — нарастать (ввиду их возможности — накапливаться в биосфере). Появилась проблема защиты организмов от ксенобиотиков и здесь на выручку многоклеточным организмам, очевидно, приходят микроорганизмы, располагающие не только системой цитохромов, использующих 02 В процессах энергетики но и в процессах пластики и детоксикационных процессах, к которым относится цитохром Р-450 и вся система оксигеназ смешанных функций (ОСФ).
Эта система располагается в цитоплазме клеток многих органов и тканей (более всего в печени, в значительно меньшей степени в легких, кожи, некоторых клетках крови, почках, головном мозге и др.). Она служит для обезвреживания попадающих в организм вредных чужеродных веществ — ксенобиотиков, а также не нужных в организме продуктов распада разрушенных тканей и клеток, излишних биологически активных веществ и продуктов их обмена – эубиотиков.
1.5 ПРИМЕРЫ ВЛИЯНИЯ АМИНОКИСЛОТ-АДАПТОГЕНОВ НА МЕХАНИЗМЫ СТРЕСС – РЕАКЦИИ
(по материалам проведенных нами ранее обзора литературы и собственных исследований: Павлов В.А. 2000.)
Как уже отмечалось выше, нормальное течение окислительно-восстановительных процессов гарантируется достаточно мощной антиоксидантной системой. Это связано с тем, во всех процессах в организме, где используется 02, образуются «побочные» продукты этих реакций — АФК — супероксидный радикал, синглетный кислород, супероксидный анион, перекись водорода, которые атакуют окружающие мембраны. В пределах физиологических изменений в организме перечисленные агрессивные вещества обезвреживаются антиоксидантной системой, в которую входят ферменты: супероксиддисмутаза, каталаза, пероксидаза; субстраты — глутатион, витамины А, Е, С, простагландины, глюкокортикоидные, адреналовые гормоны и ряд других веществ. В связи с этим ее можно рассматривать как многоступенчатую защиту тканей.
При достаточной мощности антиоксидантной системы обеспечивается необходимая интенсивность окислительных процессов, а, следовательно, и приемлемый в данных условиях уровень адаптивных возможностей организма. Воздействия, превышающие защитные возможности антиоксидантной системы, сопряжены с повреждением клетки, тканей и организма в целом.
Другим важнейшим условием интенсивного протекания окислительных процессов является наличие в достаточном количестве соответствующих субстратов. Субстратами микросомального окисления являются жирорастворимые соединения, поступающие в организм извне, чаще имеющие в своем составе циклические структуры (производные бензола, ПАУ и т.п.), а также эндогенные жирорастворимые вещества: стероидные, адреналовые гормоны, продукты деградации гемоглобина и т.д. Имеются сведения о том, что отсутствие субстратов микросомального окисления значительно увеличивает уровень АФК в них. Добавление in vitro, например, бенз(а)пирена снижает уровень АФК, следовательно, в роли антиоксидантов могут быть и канцерогенные вещества. Решающее значение в реализации адаптивных возможностей организма имеет не емкость окислительных систем (первая фаза детоксикации), а мощность ферментативных систем второй фазы детоксикации и ее субстратное обеспечение. Увеличение детоксикационных возможностей организма повышает его адаптивные возможности.
Процесс микросомального окисления рассматривается с позиций его детоксицирующего значения для организма, поскольку вслед за внедрением атомарного кислорода в субстрат в микросомах происходит конъюгация активированных соединений с глутатионом, глюкуроновой кислотой, фосфоаденозинфосфосульфатом, аминокислотами: глицином, глутамином, таурином — и затем выведение из организма.
Образующиеся в надпочечниках, щитовидной железе соответствующие гормоны могут играть роль модуляторов описанных выше механизмов.
Ключевую роль в процессах конъюгации занимает глутатион, трипептид, состоящий из глутаминовой кислоты, глицина и цистеина. Одновременно глутатион важен для антиоксидантной системы, обеспечивающей необходимый уровень протекания процессов окисления вообще. Продолжительное повреждающее воздействие на организм сопровождается накоплением адреналовых и стероидных стрессовых гормонов, что так же, как и поступление ксенобиотиков может привести к чрезмерной генерации свободных радикалов и истощение субстратов конъюгации, прежде всего глутатиона.
В условиях резкой активации микросомального окисления, усиления ПОЛ, в митохондриях нарушается работа дыхательной цепи, происходит разобщение окисления и фосфорилирования. В этих условиях митохондрии набухают, окислительные процессы в них сопряжены не с образованием энергии АТФ, а с наработкой глутаминовой кислоты. Окислительное фосфорилирование при этом угнетается, стимулируется гликолиз, происходит закисление цитоплазмы и лабилизация мембран лизосом. Последнее определяет появление в клетке активных протеин аз и расщепление белков, что способствует увеличению в тканях свободных аминокислот. Все эти изменения создают условия для стимуляции пролиферативных процессов и одновременно для усиления антиокислительной защиты клетки.
Более чем для 50 распространенных патологических состояний доказано ведущее значение в их патогенезе нарушения антиоксидантных возможностей организма.
Сюда можно отнести атеросклероз, опухолевый рост, различные воспалительные и инфекционные процессы как бактериальной, так и вирусной этиологии, в том числе туберкулёз.
Показано, что ферментные и субстратные системы, ответственные за защиту клетки от АФК эффективны лишь в интеграции с системой глутатиона. Она является последним и наиболее важным звеном в защите клетки от окислительного стресса.
Возможность глутатиона выполнять антиоксидантную и детоксикационную функцию модулируется наличием других антиоксидантных субстратов: витаминов А, С, Е, липоевой кислоты. В эксперименте установлено, что глутатион и П-токоферол представляют некую интегральную систему, в которой более стабильное значение имеет первый.
Повышенное количество АФК в ткани приводит к быстрому истощению антиоксидантных возможностей токоферола, тогда как глутатион поддерживается в восстановленном состоянии за счёт работы пентозного цикла и малик-фермента, восстанавливающих НАДФ.
Установлена также тесная взаимосвязь восстанавливающей и антиоксидантной активности глутатиона с обменом аскорбиновой кислоты. Последняя поддерживает глутатион в восстановленном состоянии.
Имеется также зависимость антиоксидантной и детоксикационной ёмкости глутатионовой системы от наличия в ткани или инкубационной среде витамина А, который усиливает глутатионпероксидазную активность, а также активность глутатион-S-трансферазы при воздействии на организм ксенобиотиков.
При чрезмерной нагрузке свободными радикалами антиоксидантные возможности атокоферола, аскорбиновой кислоты, каратина, простаглондинов исчерпывается и в случае, если система глутатиона угнетается, это сопровождается нарушением биосинтеза белка и повреждением клетки с последующим повреждением мембран, активизацией гидролитических ферментов лизисом, нарушением митохондриального окисления, деградацией метаболизма и гибелью клетки, что имеет место, как уже отмечалось выше, при интенсивном воздействии ксенобиотиков и туберкулезном поражении ткани.
Имея ведущее значение в процессе конъюгации и выведения обезвреживаемых ароматических веществ из организма, глутатион имеет не меньшее значение в механизмах антиоксидантной защиты клетки, нейтрализуя АФК и другие реакционные метаболиты: гидроперекиси липидов, повреждённые компоненты нуклеиновых кислот. И если в первом случае вступление глутатиона в реакции конъюгации обеспечивается соответствующими глутатион-S-трансферазами, то во втором — глутатион-пероксидазами.
Ферменты, обеспечивающие конъюгацию выводимых из организма ксенобиотиков, соответствующие трансферазы. Это ферменты обеспечивающие связывание активированных метаболитов с глутатионом, глюкуроновой кислотой, фосфоаденозинфосфосульфатом и аминокислотами: глицином, глутамином, таурином, цистеином.
В экспериментах установлено, что ферменты, обеспечивающие детоксикацию ксенобиотиков, обладают чрезвычайным полиморфизмом. Так, только для цитохрома Р-450 известно несколько десятков изо форм. Это же самое можно сказать о ферментах конъюгации — глутатион-Sтрансферазе, имеющей три изоформы и обладающей к тому же пероксидазной активностью и ответственной за глутатионзависимую защиту от ПОЛ, то же самое можно сказать об изоферментах уридиндифосфоглюкуронилтрансферазы. К этому следует добавить чрезвычайный полиморфизм активности ферментов у различных видов животных по отношению к различным субстратам. Имеются видовые отличия в обеспеченности субстратами, необходимыми для конъюгации: глутатионом, глюкуроновой кислотой, аминокислотами.
Введение экспериментальным животным глутатиона или компонентов, из которых состоит этот трипептид: цисте и на, глутаминовой кислоты, — частично или полностью предотвращает развитие окислительного стресса под воздействием ксенобиотиков.
Следует подчеркнуть, что защитный эффект отмеченных веществ выражен в большей степени, чем других антиоксидантов. Положительный эффект от введения перечисленных аминокислот был отмечен при различных интоксикациях, воздействии радиации, гипоксии различного генеза.
Накапливается всё больше фактов, что глутатион фокусирует в себе не только антиоксидантные и детоксикационные ресурсы организма: аминокислоты, входящие в этот трипептид, входят в важнейшие метаболические циклы и регуляторные системы различного гомеостатического уровня. Так, биосинтез глутатиона сопряжён с потреблением большого количества 5-0ксипролина, источником которого может быть соединительная ткань .
Следовательно, обмен глутатиона в определённой степени связан с обменом соединительной ткани, нарабатывающей большие количества пролина и принимающей активное участие в детоксикации ксенобиотиков .
Наиболее интенсивно биосинтез глутатиона происходит в печени, откуда он перераспределяется в другие органы. Введение аминокислот-предшественников глутатиона (глутамата и цистеина) интенсифицирует этот процесс. Определенные количества глутатиона образуются и в других органах: лёгкие, почки, лимфоидная, кроветворная ткани и др. Возможно перераспределение глутатиона как между органами, так и между клетками .
Глутаминовая кислота, являясь составной частью глутатиона, в большом количестве может образовываться из оксипролина, то есть иметь большие ресурсы в соединительной ткани .
То есть, помимо глутатиона в неспецифических механизмах защиты к повреждающему воздействию большое значение имеют аминокислоты, входящие в этот трипептид. Наибольшие изменения в обмене свободных аминокислот при действии неблагоприятных факторов на организм связаны с системой глутатиона. Так, воздействие на крыс пропанола сначала резко изменяет содержание аминокислот в тканях, а затем по мере адаптивных перестроек их уровень нормализуется. Сходные результаты получены в экспериментах по воздействию кровопотери, травматического шока, алкогольной интоксикации. При экстремальных воздействиях глутаминовая кислота метаболизируются в адаптогены антиоксиданты: ГАМК, ГОМК, янтарную кислоту, аргинин; цистеин, окисляясь и декарбоксилируясь, превращается в антиоксидант и стабилизатор мембран таурин. При этом метаболиты глутамата, метионина и цистеина реализуют свои антиоксидантные и антистрессорные эффекты как на центральном, так и на периферическом уровне. Эндокринная система мобилизует большое количество катехоламинов и глюкокортикоидов, вследствие чего катаболическим воздействиям подвергаются лимфоидная система и многие белки организма с мобилизацией большого количества аминокислот, влияя на состояние нервной, эндокринной, иммунной систем, периферических тканей и интегрируя таким образом различные уровни адаптивных перестроек.
Следовательно, такие регуляторные метаболиты как аминокислоты — могут являться не только пластическим материалом для биосинтеза различных белков организма, определяющих его специфичность, но и, включаясь в регуляторные и антиоксидантные системы, определять его неспецифическую резистентность.
В нейротрофической регуляции организма принимают активное участие гормоны и другие биологические активные метаболиты. Накапливается все больше фактов, что аминокислоты, то есть, метаболиты, являющиеся родоначальником живой материи, и мобилизующиеся в организме наряду с другими регуляторными метаболитами при повреждающих воздействиях на организм также обладают важным регуляторным воздействием. Так, в ЦНС образуется большое количество глутамата, глицина и серосодержащих метаболитов, способных оказывать мощное адаптивное воздействие при стрессовых состояниях.
Так, глутаминовая кислота обладает чрезвычайно широкой биологической активностью и имеет важнейшее значение в адаптации организма к повреждающим воздействиям. Прежде всего, сопрягая энергетический обмен с пластическим. В то же время глутамат является иммуномодулятором, стимулирующим лимфоидную систему организма .
Глутаминовая кислота также является нейромедиатором, выполняющим возбуждающую функцию в центральной нервной системе. Но в условиях стрессового воздействия на организм глутамат в ЦНС и других тканях организма интенсивно метаболизируется в гаммааминомасляную кислоту (ГАМК) — мощный тормозный медиатор и антистрессовый метаболит. В процесс е превращения глутамата в ГАМК образуется янтарный полуальдегид, обладающий антиоксидантными свойствами.
Установлено, что применение глутаминовой кислоты и её солей (натриевой и кальциевой) увеличивает концентрацию глутатиона и антиоксидантные возможности организма при высотной, гемической гипоксии, алкогольной интоксикации.
Глутамат нормализует нарушенные функции эндокринной системы (щитовидной железы, надпочечников, поджелудочной железы).
Глутаминовая кислота является фармакопейным препаратом, широко используемым в качестве средства, нормализующего тяжёлые нарушения ЦНС, а также препарата, снижающего повреждающее воздействие противотуберкулёзных препаратов на печень, что также связано с усилением биосинтеза глутатиона.
Кроме того, глутамат оказывает нормализующее воздействие на функцию митохондрий при экстремальных воздействиях на организм, стимулирует биосинтез мочевины при этих воздействиях, способствуя нормализации кислотно-щелочного состояния организма. Глутамат натрия стимулирует костномозговое кроветворение при постгеморрагической анемии. Глутамат натрия модулирует экспрессию цитохрома Р-450 через выработку гормона роста.
Изложенное позволяет сделать заключение, что глутаминовая кислота может рассматриваться как адаптоген широкого спектра действия и изменения в её обмене тесно связаны с изменением уровня глутатиона в тканях, а следовательно, антиоксидантными и детоксикационными ресурсами.
Второй аминокислотой, входящей в глутатион, является глицин, так же, как иглутамат, осуществляющий нейромедиаторную функцию в ЦНС. Глицин также как иглутамат используется при ряде патологических состояний в качестве адаптогена.
И, наконец, третьим и последним компонентом глутатиона является цистеин — серосодержащая аминокислота. Цистеин может образовываться только из метионина незаменимой аминокислоты. Превращение в цистеин возможно лишь после активного включения метионина в процессы метилирования, например, фосфолипидов, креатина, пиримидиновых оснований и других метаболитов. То есть, количество цистеина в тканях зависит от интенсивности включения в обмен метионина. Цистеин обладает антиоксидантной активностью.
Глутатионовая конъюгация завершается образованием конъюгатов меркаптуровых кислот, представляющих собой остаток цистеина и процесс обезвреживания эндогенных токсинов и ксенобиотиков сопряжён с выведением из организма большого количества незаменимых серосодержащих аминокислот. Для обеспечения антиоксидантных и детоксикационных процессов должен быть высокий уровень обмена метионина и необходимое поступление его с пищей. В этом плане интересно отметить, что у морских свинок, неспособных синтезировать аскорбиновую кислоту, так же, как и у человека, уровень серосодержащих аминокислот (метионина, цистеина, таурина) в большинстве тканей ниже, а глутатиона, особенно восстановленной его формы, выше, чем у животных, синтезирующих витамин С.
Аминокислоты, входящие в глутатион, могут быть предшественниками других мощных антиоксидантных субстратов, нежели глутатион. Так, глутаминовая кислота является мощным стимулятором биосинтеза аргинина и мочевины в печени, являющихся антиоксидантами. Метаболизируясь в орнитин, глутаминовая кислота служит предшественником полиаминов. Аргинин, мочевина и полиамины обладают мощным антиоксидантным потенциалом. Эти вещества накапливаются в тканях при экстремальных воздействиях на организм — низкая температура, повышенное давление 02, радиация.
Важно отметить, что метионин, активируясь и превращаясь в S-аденозилметионин, включается в процесс биосинтеза полиаминов при резком снижении в тканях глутатиона и увеличении АФК, стимулирующих процессы декарбоксилирования.
В первом случае это включение глутамата и серосодержащих аминокислот в глутатион и — во втором — включение их в полиамины, при увеличении биосинтеза аргинина и мочевины в печени, которые стимулируются глутаматом и глутамином. Второй путь преобладает при истощении системы глутатиона и усилении свободнорадикальных процессов в клетке. Именно свободнорадикальному механизму отводится решающая роль в канцерогенном воздействии ксенобиотиков и других неблагоприятных факторов на организм. Возможно, что в данной ситуации организм, исчерпав возможности адекватной физиологическому уровню антиоксидантной и детоксикационной защиты с преобладанием системы глутатиона, переходит на другие защитные механизмы, связанные с образованием большого количества азотистых антиоксидантов: аргинина, мочевины, полиаминов.
Решающее значение в такой перестройке может иметь метаболический фонд свободных аминокислот, прежде всего глутаминовой и серосодеожащих (метионин и цистеин), и уровень АФК и других радикалов в ткани — влияющих на интенсивность декарбоксилирования.
Представленные механизмы явились основанием для использования глутатиона и серосодержащих аминокислот в качестве веществ, защищающих организм от ксенобиотиков, радиации и ряда других повреждающих воздействий. В то же время при таких воздействиях как повышенное давление кислорода, пониженная температура, защитного эффекта удаётся достичь, используя аргинин, мочевину, то есть, богатые азотом, но не содержащие серу аминокислоты и их производные.
Имеются экспериментальные данные о том, что рассматриваемые аминокислоты могут влиять на состояние организма через модуляцию его иммунной системы и ЦНС.
Содержание свободных аминокислот и их производных высокостабильно для различных тканей организма. Поэтому изменение этих параметров может существенно изменить метаболический фонд важнейших обменных процессов в клетке .
Мощным источником глутаминовой кислоты и глицина в организме может быть ЦНС и СТ. Усиленное включение в биосинтез так, при болезни Альцгеймера нарушение образования в головном мозге глутамата, глицина и некоторых других аминокислот приводит к нарушениям азотистого обмена во всём организме и его адаптивных возможностей. О том, что значительные количества глутамата и глицина образуются в мозговой ткани, свидетельствуют и другие источники.
Всё изложенное свидетельствует о том, что последствия повреждающих воздействий на организм во многом зависят от его способностей мобилизовать в достаточной степени субстраты, необходимые для эффективного функционирования системы глутатиона, то есть соответствующие аминокислоты.
1.6 СТРЕСС ВОЗДЕЙСТВИЕ, КАК ФАКТОР СТИМУЛЯЦИИ И ИНТЕГРАЦИИ ЗАЩИТНО-АДАПТИВНЫХ СИСТЕМ ОРГАНИЗМА, ПОПУЛЯЦИИ И ОБЩЕСТВА.
Многочисленные исследования, посвященные стресс воздействию на организм, позволили сформировать учение о стрессе, как универсальной реакции организмов на повреждение и, как позднее выяснилось, вообще всей живой материи на внешние воздействия. Однако реакция эта становится все более сложной и эффективной по мере филогенетического совершенствования живой материи и смысл ее состоит в повышении адаптивных возможностей организмов, приспосабливающихся к изменениям окружающей среды. У высших, в функциональном плане, организмов-млекопитающих стресс-реакция наиболее сложная и состоит в ответе защитно-адаптивных систем организма на различные воздействия и закреплении приобретенных свойств устойчивости к неблагоприятным воздействиям на генетически-метаболическом уровне. Таким образом, стресс-воздействие и адаптивный ответ на него являются единой системой, обеспечивающей функциональное развитие живой материи.
Имеются многочисленные сведения, что воздействия, предъявляющие повышенные требования к адаптивным системам организма одновременно стимулируют их, что является важнейшим механизмом эволюционного развития (Яковлев Г.М. и др. 1990, Судаков К.В. 1984, 1997, Самюэль К 2000, Менделевич В.Г. 2002). Проведенные нами исследования позволили установить, что различные стресс-воздействия по-разному влияют на различные виды животных, располагающих различными защитно-адаптивными ресурсами, и особенностями специфического и неспецифического ответа на внешние раздражители. Ведущее значение в этих различиях имеют соотношения в тканях и клетках адаптивных систем аминокислот – адаптогенов. В зависимости от ресурса в организме обогащенных азотом или серой аминокислот можно выделить 2 крайних защитно-адаптивных типа – N- и S-тип.
Здесь необходимо дать краткую характеристику процессов совершенствования адаптивных механизмов живой материи по мере эволюционного развития филогенеза.
На сегодня установлено, что жизнь на планете Земля возникла около 3 млрд. лет назад.
Согласно теории Опарина-Холдейна в определенных физико-химических условиях, имевших место на планете в то время, из неорганических веществ (Н2, СО2, NH3, Н2О и ряда других) образовались новые органические молекулы – некоторые аминокислоты, липиды и ряд других.
В лабораторных условиях, впервые Миллеру, а затем другим исследователям удалось получить из перечисленных выше неорганических веществ 4 аминокислоты (глицин, аланин, аспарагиновая и глутаминовая аминокислоты, являющиеся родоначальниками других аминокислот), липиды и ряд других органических молекул, способных в дальнейшем трансформироваться и взаимодействовать, давая другие органические молекулы. Эти опыты являлись убедительным аргументом в пользу теории Опарина-Холдейна.
Зарождение и первые этапы жизни на Земле проходили в анаэробных условиях. Более 2-х млрд. лет господствующей формой жизни на планете были прокариоты – микроорганизмы, во многом предопределившие как основные направления развития живой материи, так и важнейшие процессы геохимии Земли (Шлегель Г. 1987).
Революционное значение в процессах развития живой материи имело появление в водной среде океанов пурпурных зеленых бактерий, а затем цианобактерий – сине-зеленых водорослей, способных к фотосинтезу и образованию О2. Процессы фотосинтеза у бактерий и растений обеспечивали и обеспечивают до сих пор биосферу Земли как органическим материалом так и О2. И то и другое необходимо для непрерывного совершенствования адаптивных механизмов представителей животного мира и приспособления к имеющимся факторам окружающей среды (Шлегель Г, 1981).
Однако уровень О2 в атмосфере как и структура биомассы Земли существенно менялись в различные временные периоды. Так, первоначально концентрация О2 была довольно низка – не более 2% вплоть до периода 0,6-0,5 млрд. лет назад. Но по мере того как фотосинтезирующие растения сплошь покрыли все материки концентрация О2 непрерывно возрастала и составила около 100-80 млн. лет назад 60-40% состава атмосферного воздуха (Наука и жизнь, №6 1985).
В образцах воздуха из пузырьков янтаря и древних отложений льда того периода обнаружена именно такая концентрация О2.
Аналогичная концепция о существенном повышении концентрации кислорода во времена каменноугольного периода высказывалась учеными Йельского университета и Американского геологического общества Робертом Бернером и Гарри Линдисом. Значительные концентрации кислорода приводили к интенсивным пожарам и существенно повышали плотность атмосферного воздуха, что делало возможным полет гигантских рептилий и насекомых.
В связи с этим дальнейшая эволюция живой материи потребовала более сложных генетически – метаболических форм и соответственно функционально более совершенных форм организации, нежели прокариоты и растительный мир. Появляются эукариоты – в подавляющем большинстве аэробные организмы. Сначала средой их обитания была водная стихия океанов. Затем они вышли на сушу и распространялись по новым материкам планеты в виде земноводных и пресмыкающихся.
Обращает на себя внимание чрезвычайный полиморфизм и огромная масса последних. Это могло быть связано с несколькими факторами.
300-100 млн. лет назад высокая концентрация О2 в атмосфере неизменно приводила к интенсивным пожарам– сгоранию и перегонке обильно разросшегося растительного мира в результате чего образовывались биологически активные вещества (БАВ) – прежде всего полициклических ароматических углеводородов (ПАУ), накопившиеся в биосфере. (Необходимо отметить, что нефть и уголь, образовавшиеся много миллионов лет назад, содержат большое количество ПАУ). ПАУ – жирорастворимые вещества, легко проникающие через биологические мембраны, резко изменяющие их свойства вплоть до повреждения и разрушения (Нечипоренко С.П., 1981; Голиков С.И., 1988).
Центральным звеном обеспечения жизнедеятельности всех живых организмов являются окислительно-восстановительные реакции, но высокие концентрации О2 (сильный окислитель, повреждающий клетки живой материи) и ПАУ поставили взаимодействующую с ними живую материю перед выбором – либо погибнуть или уйти в недоступные для этих веществ ниши – например, глубины океанов, что некоторые представители животного мира и сделали, либо перейти на новый метаболически-адаптивный уровень. И этот переход состоялся, породив новые, гораздо более перспективные в адаптивном плане, чем прокариоты и представители флоры формы жизни – эукариот, представителей фауны.
Развитие животного мира сосредоточилось на совершенствовании аэробных эукариат. – занявших доминирующее положение среди фауны. Использование О2 в качестве окислителя в ОВП резко повысило функционально-адаптивные возможности новых видов и одновременно создало идеальные условия для интенсивного процесса естественного отбора в жестких условиях гипероксидативной среды насыщенной мутагенными БАВ – ПАУ.
С этих позиций становится тоже понятными причины огромного разнообразия обнаруженных ископаемых пресмыкающихся в соответствующие периоды развития биосферы Земли (100-50 млн. лет назад).
Основываясь на данных современной токсикологии можно попытаться реконструировать адаптивно-метаболические процессы и их совершенствование у животных того времени.
Важнейшее значение в защите важнейших структур клеток и тканей от агрессивных метаболитов при метаболизировании их в тканях являются система детоксикации, протекающая в 2 фазы в эндоплазматической сети клетки.
I фаза – окисление проникших в клетку ПАУ в системе ОСФ, чтобы в дальнейшем они могли быть связаны со специальными обезвреживающими веществами и выведены из клеток и тканей организма через почки, легкие, кожу.
II фаза – процесс связывания токсичных веществ, в том числе и продуктов обмена (БАВ), с некоторыми специфичными, предназначенными для этого веществами (УДФГК, ФАФС, глутатион, некоторые аминокислоты – глицин, глутамин, таурин). Особенно большое значение в процессах обезвреживания ПАУ и промежуточных продуктов обмена О2 имеет глутатион (Нечипоренко С.П., 1981; Голиков С.И., 1988).
Однако в условиях перенасыщенности среды О2 и ПАУ отмеченных механизмов для их обезвреживания недостаточно (Кричевская А.А., 1981). Необходимы мощные вещества — адаптогены, защищающие прежде всего от повреждения мембраны. К таким веществам относятся: таурин, полиамины, аргинин, мочевина, нуклеиновые кислоты, мочевая кислота и некоторые другие вещества, образующиеся прежде всего в печени и оказывающие мощное мембраностабилизирующее действие (Кричевская А.А., 1981, 1982; Шейбак Л.Н., 1996; Нефедов Л.И., 1992, 1999). Одновременно все эти метаболиты оказывают стимулирующий эффект на процессы пролиферации и роста. Возможно, именно эти обстоятельства и определили огромную массу динозавров – как своеобразную защиту к избытку О2 и ПАУ. В то же время высокие мутагенные свойства ПАУ и АФК определяли высокий уровень мутаций, многообразие видов и невысокие адаптивные возможности возникающих видов.
Строго говоря, теория Дарвина, которая связывает естественный отбор с генетическими мутациями, справедлива именно для этого периода развития живой материи. Однако с исчезновением динозавров и появлением новых доминирующих классов животных – млекопитающих значение генетических мутаций в процессах совершенствования защитно-адаптивных механизмов существенно снижается. Как на сегодня признанно большинством специалистов в этих областях научного знания, для млекопитающих большее значение имеет трансгенный перенос, осуществляемый микроорганизмами и размах генетических и фенотипических изменений существенно ниже, чем в предшествующую эпоху (Л. Маргулис, 1998; П.Л. Луизи, 1993).
Поэтому для млекопитающих можно говорить не о генетически-метоболических ЗАМ, а скорее об адаптивно-метаболических, в которых ведущее значение имеет не значительное изменение генетического аппарата, а изменение метаболических взаимоотношений. В этих модификациях большое значение имеет взаимодействующая с макроорганизмом микрофлора, а не высокие концентрации агрессивных и мутагенных веществ. Поэтому следующим этапом совершенствования адаптивных механизмов после генетически-мутационного можно назвать этапом метаболической компартментализации или метаболически-адаптивный этап.
Снижение концентрации О2 в атмосфере при сохранении в ней большого количества ПАУ (они накапливаются в биосфере и могут долго сохраняться) вследствие массовой гибели реликтовых папоротниковых лесов сделали нежизнеспособными гигантских О2 и ПАУ зависимых пресмыкающихся. Доминирующее значение стали приобретать животные, способные эффективно утилизировать О2 и перераспределять его прежде всего для нужд нервной системы – мозга и одновременно способные сохранять устойчивость метаболических процессов и повреждающему воздействию ПАУ. Своеобразным реликтовым видом, обладающим такими возможностями, является крыса (Ж. Сен — .Леран 1968).
По мнению многих специалистов именно на крысу по многим параметрам походили первые млекопитающие.
И действительно, особенности обмена веществ у крысы таковы, что они занимают как бы промежуточное положение в плане защитно-адаптивных ресурсов и механизмов между животными, жившими в гипероксидативных условиях и высокой концентрации ПАУ и животными другой эпохи – живущих при гораздо более мягких в токсикологическом плане условиях.
Особенности обмена веществ у крысы состоят в том, что в защитно-адаптивных механизмах отмечается высокая чувствительность к воздействию глюкокортикоидов, мутагенных и канцерогенных веществ, прежде всего ПАУ (Шабад Л.М 1970, Шапот В.С 1975, Голиков П.И., 1985).
Имеются многочисленные работы в которых показано, что ГК претерпевают те же обменные превращения, что и ПАУ с воздействием продуктов их обмена на генетический аппарат клетки, а так же на многие другие рецепторные, мембранные, ферментные и другие регуляторные центры и метаболические процессы (Нечипоренко С.П., 1982; Голиков С.И., 1986).
Основой структуры стероидных гормонов, в том числе и ГК, витаминов группы Д, холестерина является молекула циклопентанпергидрофенантрена, то есть ПАУ. Все это наводит на мысль о том, что перечисленные БАВ – это своеобразная интернализация ПАУ — превращение их из экзогормонов среды в эндогормоны организма. В подтверждение этой гипотезы можно привести то обстоятельство, что обезвреживание ПАУ в организме начинается с их окисления при участии цитохрома Р-450 и образование ГК в надпочечниках также происходит с использованием биосинтезирующим аппаратом их клеток цитохрома Р-450 (Голиков П.П. 1988).
Реализации эффектов ГК при стресс-воздействии посвящена большая литература. Основным механизмом воздействия считается стимулирование протеолитических ферментов в иммунной, соединительной, мышечной и ряде других тканей организма. Однако многие механизмы ГК воздействия во многом так и не раскрыты, установлено лишь, что имеются некие белково-рецепторные структуры внутри клеток (Rp I, II, III) с которыми связываются ГК. Считается что ГК влияют на генетический аппарат клеток, снимая репрессию со структурных генов и усиливая наработку соответствующих белков-ферментов, кроме того доказано активизирующее влияние ГК на различные ферменты, например тирозин-аминотрансферазу, триптофандиоксигеназу, ферменты глюконеогенеза и ряд других. С другой стороны ГК ингибирует некоторые ферментные системы, например ферменты гликолиза и цикла Кребса.
Несомненным также является тот факт, что ГК, подвергаясь обмену в тканях, способствует генерации большого количества АФК и реакционоспособных метаболитов ПАУ (Голиков П.П., 1988).
Повышение количества АФК обеспечивает прямое окисление СЖК при стресс-реакции при повышенном уровне их в организме, с образованием необходимых количеств АТФ для реализации ЗАМ в условиях подавления гликолиза и ЦТК.
Именно при таком типе ответа на стресс-воздействия организм более устойчив к острым и интенсивным повреждающим факторам, что мы и наблюдаем при воздействии таких факторов на крысу.
Необходимо уточнить, что основные детексикационные процессы с участием цитохрома Р-450 по метаболизированию ПАУ, ГК и КХА а также многих других ксено- и эубиотиков протекают преимущественно в печени.
На основании полученных результатов исследований и анализа литературных данных мы сочли возможным составить следующую таблицу:
Периодизация филогенеза и адаптивных возможностей живой материи в зависимости от изменений концентрации кислорода в атмосфере:
1 – Физико-химический этап (образование органических молекул) — бескислородный — 4-3 млрд. лет назад;
2 – Царство прокариот — бескислородный — 3-2,5 млрд. лет назад;
3 – Фотосинтетический период (прокариоты и фотосинтезирующая флора) — накопление кислорода (2%) — 2,5-0,5 млрд. лет назад;
4 – Генетический мутационный (преобладание земноводных и гигантских пресмыкающихся)- максимальное накопление кислорода (40-60%) — 300-50 млн. лет назад;
5 – Метаболически-адаптационный (формирование адаптивных центров – печени и мозга – период цефализации) — горение первичных лесов, накопление в атмосфере БАВ-ПАУ, снижение концентрации кислорода — 50-2 млн. лет назад;
6 – Метаболически-коммуникативный (появление и развитие человеческого общества, снижение адаптивности флоры и фауны) — снижение концентрации кислорода, ПАУ — 100 тыс. лет назад до настоящего времени;
7 – Ноосфера — сознательное поддержание адаптивности человека и окружающей природы — будущее развитие человека и биосферы.
1.7 МОЗГ И ПЕЧЕНЬ, КАК ДВА ОСНОВНЫХ ПОЛЮСА ОБРАЗОВАНИЯ АДАПТОГЕНОВ ОРГАНИЗМА В ЭВОЛЮЦИОННОМ АСПЕКТЕ.
Выдвигаемая нами концепция о различных защитно-адаптивных типах по сути дела является развитием положения о цефализации как о важнейшем механизме эволюционного развития живой материи.
Исходя из наших данных, главенствующее значение в совершенствовании ЗАМ имеет метаболическая функция ЦНС и прежде всего мозга в интеграции с другими защитно-адаптивными системами, прежде всего с печенью, соединительной тканью, системой кроветворения и иммунитета.
При этом ЦНС и мозг играют не только координирующую роль в жизнедеятельности организма, но имеют самостоятельное значение в образовании, накоплении и мобилизации важнейших метаболитов-адаптогенов организма в условиях ответа на стресс-воздействие. Это во многом и предопределяет реализацию и совершенствование ЗАМ и формирование более адаптивных защитно-адаптивных типов.
Особенно большое значение мозг как орган образования адаптогенов имеет при эмоциональном стрессе (ЭС).
Еще раз вкратце напомним, что классические представления о стрессе – это смена фаз: тревоги — резистентности – истощения, во многом предопределяющаяся изменениями гормонально-метаболического фона. Инициируется процесс повышенным образованием в ЦНС и надпочечниках КХА с выбросом их в кровь, стимулирующих ферменты мобилизации из депо углеводов и жиров со значительным повышением их в крови. Это повышение сопровождается усиленным образованием в клетках поджелудочной железы инсулина, обеспечивающего метаболическую утилизацию повышенных количеств углеводов и жиров из крови тканями защитно-адаптивных систем: сердечно-сосудистой, дыхательной, кроветворной и другими, активно участвующими в стресс-реакции. Усиление в тканях ОВП с образованием энергии АТФ позволяет существенно повысить их функциональную активность и сопротивляемость к стресс-воздействию. Однако эти процессы, определяемые воздействием повышенных количеств КХА кратковременны. На смену им идет гормонально-метаболический фон, определяемый образованием повышенных количеств ГК, который собственно и обуславливает основные адаптивные сдвиги в ответ на стрессовый фактор, что было установлено, в основном, на крысах. Прежде всего, происходит резкая активизация лизосомального аппарата клетки и усиление протеолиза белков лимфоидной, мышечной, соединительной и ряда других тканей. Имеются сведения, что ГК (также как и КХА), подвергаясь процессам обезвреживания (при значительном повышении их количества в различных тканях организма — печень, легкие и др.), способствуют повышению уровня АФК и свободных радикалов в форме промежуточных метаболитов обезвреживания КХА и ГК (Нечипоренко С.И., 1981, Голиков С.И 1984, Меерзон Ф.З. 1984, 1999). Это во многом и предопределяет усиление протеолических процессов. Происходит расщепление белков в тканях (прежде всего в лимфоидной, соединительной, мышечной ) в том числе и многих рецепторных белков как мембранных, так и внутриклеточных (например, рецепторов к ГК – II-III типа) (Голиков П.П. 1988, Кульберг А.Я. 1987, 1988), что сопровождается значительным повышением свободных аминокислот в тканях организма. Ряд аминокислот являются мощными адаптогенами. Так, аланин, глутамин, аргинин, аминокислоты с развлетвленной углеродной цепью являются стимуляторами анаболических процессов (Кричевская А.А. 1981, Козлов Н.Б. 1971). Другие аминокислоты обладают антиоксидантным эффектом (глутамат, аргинин, цистеин, таурин). 10 аминокислот стимулируют процессы глюконеогенеза в печени и почках, восполняя тем самым ресурсы углеводов в организме, подвергающимся стресс-воздействию (Кендыш И.Н. 1985). Все эти механизмы стабилизации метаболических процессов в условиях воздействия повреждающего фактора описываются понятием реконструктивный протеолиз и направлены на выход организма на новый метаболический уровень – более адекватный повреждающему воздействию (Покровский А.А. 1986).
Мы считаем, что особенности стресс – реакции и адаптивного ответа на нее определяются различиями на уровне регуляторных метаболических систем, ведущее значение в которых имеют аминокислоты адаптогены. Их ресурс, наследуемые и изменяющиеся в процессе жизнедеятельности организма соотношения.
На то, что стресс-воздействия могут быть разными как по силе, так и по степени и особенностям адаптивного ответа организма указывал еще Г. Селье (1981 г.). Необходимо подчеркнуть, что ЭС существенно отличается от других видов стресса по своим приспособительно-метаболическим механизмам и имеет ключевое значение в механизмах эволюции живой материи вообще и в совершенствовании процессов цефализации в частности. Это особенно важно для становления человека, человеческого общества и его развития. Особенности ЭС состоят в том, что в отличие от других видов стресса, здесь имеет ведущее значение психо-эмоциональный фактор. Основными медиаторами ЭС являются КХА – производные аминокислот фенилаланина и тирозина (норадреналин, адреналин и дофамин) вырабатывающиеся в различных структурах мозга и мозговом веществе надпочечников при I фазе стресса – фазе тревоги (Судаков К.В. 1984, 1997, 2004, Черков А.Н. 1989, Раевский К.С., Георгиев В.П. 1986).
КХА мобилизируют из имеющихся в организме депо углеводы (глюкозу) и липиды (СЖК), способствуя повышенному образованию АТФ в энергетических центрах клеток в процессе их окисления и повышению функции защитно-адаптивных систем организма с повышением возможностей организма избегать повреждающее воздействие (в природных условиях это чаще всего нападение хищников) или вырабатывать психологические защитные реакции, отвлекающие от травмирующего психоэмоционального воздействия (интродукция) (Поршнев Б.Ф. 1972).
В отличие от физико-химических воздействий окружающей среды при ЭС (особенно при его хроническом воздействии) не имеет столь большого, можно даже сказать главенствующего, значения усиление наработки надпочечниками ГК с реализацией их метаболически адаптивных механизмов. Это происходит потому, что постоянное, многократно повторяющееся психо-эмоциональное воздействие обуславливает постоянное образование КХА с реализацией их рецепторно-метаболических механизмов и не дает в полной мере включиться механизмам реализации ГК-эффектов (Панин Л.Е. 1983, Голиков П.П. 1988).
При воздействии на различных животных ГК и КХА была установлена различная чувствительность к ним различных видов животных. Наиболее значительно отличались по метаболическим последствиям введения ГК крысы и морские свинки. Крысы были высокочувствительны – ГК (+), а морские свинки напротив малочувствительны к ГК (-) (Голиков П.П. 1988). То есть, классические механизмы реализации стресс реакции на метаболическом уровне под воздействием ГК более характерны для крысы, а не для морской свинки. С чем это связано авторы не разъясняют.
Полученные нами результаты позволяют понять, в чем состоят эти различия и более полно представить механизмы адаптивных перестроек в ответ на различные по силе, характеру воздействия повреждающего фактора.
В наших исследованиях установлено, что ГК (+) крысы и ГК (-) морские свинки по-разному реагируют как на ЭС, так и на другие стресс-воздействия. И эти различия связаны, прежде всего, со способностями нарабатывать аминокислоты-адаптогены и некоторые другие метаболиты, обладающие адаптогенным эффектом, в ответ на повреждающие факторы (Павлов В.А. 2000).
Кардинальное отличие в метаболизме у этих животных состоит в том, что морские свинки (также как приматы и человек) не могут синтезировать аскорбиновую кислоту (АК). Поэтому стресс-воздействие у крыс сопряжено с усиленным образованием АК и изменением обмена соединительной ткани (СТ) в сторону повышенного образования оксипролина и усиленным образованием СТ с потреблением необходимых для этого аминокислот – пролина (глутамата) и глицина с уменьшением их количества в крови (Павлов В.А. 2000).
Одновременно в печени у крыс образуются большие количества таурина, содержание которого повышается в тканях организма, ответственных за повышение его адаптивного уровня. Такой ход метаболических перестроек при более или менее длительном воздействии физического стресс-фактора (температура, радиация, физическая нагрузка, инфекции и т.д.) на организм позволяет выйти на метаболический уровень, соответствующей фазе резистентности (Павлов В.А. 2000-2004).
В классическом варианте это означает снижение интенсивности образования инсулина поджелудочной железой, при чем становится достаточным небольшое увеличение продукции ГК надпочечниками для поддержания метаболически-адаптивных механизмов стресса. При этом гормональный фон перестраивается таким образом, что адаптивные ресурсы организма направляются на восстановительно-биосинтетические процессы, стимулируемые на уровне генетического аппарата глюкокортикоидами по механизму дерепрессии генов-операторов. Это определяет усиление работы структурных генов, передающих информацию на информационную РНК, с последующим образованием на рибосомах клетки необходимых для адаптационных перестроек белков-ферментов, с усилением сопротивляемости к повреждающему воздействию. Это мы и наблюдали у крыс при воздействии на них различных видов гипоксии, токсичных доз ксенобиотиков, кровопотери, заражения МБТ, проникающей реакции. Эти же воздействия на морских свинок приводили к довольно скорой их гибели, без существенных адаптивных перестроек, но они были высокоустойчивы к мутагенным воздействиям, а также к ЭС. Что определяло столь существенные различия?
В экспериментах на животных нами было установлено, что многие метаболические процессы, имеющие отношение к адаптивным перестройкам у этих видов животных были различными (Павлов В.А. и соавторы 2000, 2001, 2002, 2004). Так, у морских свинок в тканях было в 2 раза больше железа и серы, чем у крыс. Основное количество железа и серы в тканях входит в железосерные комплексы окислительно-восстановительных ферментативных центров, определяющих интенсивность протекания ОВП в митохондриях (ферменты дыхательной цепи) и оксигенез смешанных функций в цитозоле клеток (Хьюз М 1983, Шлегель Г. 1987). В митохондриях железо-серные центры ферментов необходимы для процессов окислительного фосфорилирования и образования энергии АТФ. В цитозоле клетки ферменты с железосерными комплексами необходимы для детоксикации ксенобиотиков либо токсичных веществ, образующихся в ходе некоторых промежуточных метаболических превращений собственных веществ (эубиотиков), подвергающихся разрушению в системе ОСФ, включающих в себя цитохром Р-450.
Имеются сведения, что у крыс в большинстве тканей, особенно в печени намного больше цитохрома Р-450 и связанных с ним ферментов ОСФ, чем цитохромооксидаз или других ферментов дыхательной цепи митохондрий, необходимых для процессов окислительного фосфорилирования и включающих в себя железо и серу (Нечипоренко С.П. 1981).
Но исходя из наших данных при сниженных по сравнению с крысой детоксикационных возможностях железа и серы в тканях морских свинок намного больше, чем у крыс, следовательно, оно может находиться преимущественно в железо-серных комплексах митохондрий а не в ОСФ. Или другими словами морские свинки – это «митохондровики» ориентированные на процессы Эмбдена-Меергофа-Кребса. Тогда как крыс можно характеризовать как «ОСФ-ников – пентозников». Это становится еще более понятно, если учесть, что в условиях стресса повышенный ГК фон тормозит процессы расщепления и окисления углеводов (вследствие угнетения образования инсулина), а также процессы окислительно-восстановительного фосфорилирования с их разобщением. Энергия АТФ при этом образуется за счет прямого окисления жирных кислот, за счет стимуляции их перекисного окисления высоким фоном АФК в митохондриях (Голиков П.П. 1988). В этих же условиях стимуляция глюконеогенеза повышает уровень образования глюкозы из аминокислот, способствует стимуляции ГМФШ и наработки НАДФ Н2, необходимого для интенсивного протекания ОСФ, прежде всего в печени, а также образованию АК в СТ. Кроме того усиленное образование НАДФ Н2 изменяет процессы обмена и функцию мембран, а также функции многих систем, напрямую зависящих от состояния мембран, например гепатоцитов в печени или нейронов в мозге (Лабори А. 1971). Соответственно меняется их функционально-метаболические возможности.
Печень во многом обеспечивает функционально-метаболические изменения в организме в реализации стресс-воздействия и адаптивного ответа на него. И от ее ресурсов во многом зависит тип адаптивного ответа на стресс-воздействие.
Именно печень обеспечивает организм многими адаптогенами в условиях стресс-воздействия и адаптации крыс. В печени мобилизуются многие аминокислоты-адаптогены, транспортные белки и пептиды, серосодержащие аминокислоты, основные ресурсы Fe, Cu, Mn и других микроэлементов, здесь образуются транспортные формы липидов (ЛПОН, ЛПНП, ЛПВП) используемые в условиях стресса как энергетические ресурсы, а также как регуляторные факторы энергетических процессов в условиях стресс-воздействия. Кроме того, печень во многом ответственна за стабилизацию температурного режима организма, особенно в условиях холодового стресса (Панин Л.Е. 1983).
Здесь усиливается наработка НАДФН2 в ГМФШ и образование АК. Все это повышает адаптивные резервы организма, прежде всего его окислительно-восстановительные возможности и обеспечивает резистентность крыс к таким повреждающим факторам, как токсические дозы ПАУ, кровопотеря, высотная гипоксия, радиация, но не к хроническому эмоциональному стрессу, мутагенным и канцерогенным воздействиям.
У млекопитающих печень составляет около 4% от массы тела – это самый большой паренхиматозный орган, обеспечивающий адаптивные метаболические перестройки. Однако у приматов и человека печень составляет всего 2% от массы тела, примерно столько же, как и головной мозг (2%) (Шлегель Г. 1987).
Мозг также является крупнейшим паренхиматозным органом, способным нарабатывать мозговые адаптогены организма. Так, здесь образуются и в больших количествах присутствуют адаптогены широкого спектра действия, защищающие ткани организма при повреждающих воздействиях: глутаминовая кислота, аспаратиновая кислота, глутамин, ГАМК, ГОМК, цистатионин, цистеиновая кислота, таурин, изетеиновая кислота. В мозге образуются и другие БАВ, обладающие мощьными адаптивными эффектами: различные гаммаглутамилпептиды (например, глутатион), котехоламины, серотонин, эндорфины, энкефалины, олиго- и полипептиды, обладающие адаптогенным эффектом в условиях стресс-воздействия. . Некоторые из этих веществ выполняют медиаторные функции в различных отделах мозга (Раевский К.С., Георгиев В.П., 1986, Чирков А.М 1989, Робу А.И. 1989, Судаков К.В. 1984, 1997, 2004). Для других какая-либо определенная функция в головном мозге не установлена (карнозин, анзерин, цистатионин, изетеиновая кислота, N-ацетилглутаминовая кислота, N-ацетиласпарагиновая и др.) В то же время эти вещества обладают антиоксидантными, мембраностабилизирующими свойствами, например, в сердечной мышце, скелетной мускулатуре и других органах и тканях (Меерсон Ф.З. 1997, 1999). Таким образом, мозг является не только органом, воспринимающим и обрабатывающим сигналы из внешней среды и, затем, корректирующий организм сообразно изменяющейся ситуации, но, и наряду с печенью, являющийся мощным органом метаболической адаптации, нарабатывая БАВ-адаптогены, способные переходить через геметоэнцефалитический барьер (ГЭБ) и участвовать в стабилизации адаптивных систем организма. (Кричевская А.А. 1982, Раевский К.С., Георгиев В.П. 1986, Нефедов Л.И. 1992, 1999).
В то же время между мозгом и печенью имеется взаимодействие при реализации стресс-воздействия. Так, вещество, обладающее общим тормозным воздействием на возбудимые ткани – таурин, образуется преимущественно в печени, но имеет специальные транспортные системы в ГЭБ и мозговых структурах, концентрируется в тканях мозга, особенно в его сером веществе, существенно повышая тормозные ресурсы нейронов. (Кричевская А.А. 1982, Раевский К.С., Георгиев В.П. 1986, Нефедов Л.И. 1992, 1999).
С другой стороны глутаминовая кислота и глутамин в больших количествах образуются в мозге, особенно в условиях стресс-воздействия. В условиях стресса в тканях мозга освобождается большое количество аммиака, очень токсичного для нервной ткани. Связывание и, соответственно, обезвреживание аммиака осуществляется глутаминовой кислотой в глутаминсинтетазной реакции с образованием больших количеств глутамина. Глутамин легко проникает через ГЭБ и используется прежде всего в печени в биосинтетических процессах. Глутамат и глутамин используются в различных адаптивно-метаболических процессах организма прежде всего в печени и почках (Волков М.С. 1975, Павлов В.А. 1988, 2000).
Однако характер реализации стресс-воздействия может быть различным. Так, крысы и морские свинки существенно отличаются по чувствительности к воздействию стресс-факторов. Другие млекопитающие находятся в промежутке между 2-мя этими видами по чувствительности к ГК в ходе реализации стресс-реакции и адаптивного ответа организма.
Высокий уровень АФК в тканях у крыс стимулирует декарбоксилазы цистеиновой кислоты и орнитина и соответственно образование таких адаптогенов как таурин, полиамины прежде в печени и других тканях организма (Oguro T 1987).
Высокий уровень АФК способствует окислению АК и НАДФН2 и стимулирует тем самым ГМФШ, а усиление глюконеогенеза и реконструктивного протеолиза и значительное повышение количества в тканях свободных аминокислот обеспечивает его необходимым субстратом – глюкозой. Эти же обстоятельства способствуют усиленному включению глюкозы в биосинтез АК. Усиленное образование АК и НАДФН2 способствуют стабилизации ОВП и предотвращают окисление ГSH. Чему способствует также и высокий уровень таурина, что было показано в наших исследованиях при воздействии на крыс ксенобиотиков – ПАУ, кровопотере, заражении МБТ, радиации, эмоционального стресса. Таурин, ГSH, полиамины накапливаются в кроветворной, лимфоидной тканях обеспечивающих пластическую реконструкцию поврежденных тканей и структур и обеспечивает высокую устойчивость к повреждающим факторам (Павлов В.А. 1999, 2000).
Однако постоянно высокий уровень АФК, радикалов промежуточного метаболизма ГК (других агрессивных продуктов метаболизирующихся циклических БАВ, а также поступающих извне ПАУ или других мутагенных и канцерогенных ксенобиотиков) оказывает отчетливое мутагенное воздействие на генетический аппарат крысы, несмотря на высокий уровень метаболитов-адаптогенов в тканях. (Худолей В.В. 1993-1996, Шабад Л.М. 1970, Середенин С.Б. 1980).
Другой тип метаболических реакций на стресс-воздействие наблюдается у морской свинки, которая считается ГК (–) отрицательной. То есть ее рецепторный аппарат (к воздействию ГК) в виде белков- рецепторов (I, II, III) во многих тканях либо мало выражен, либо вообще отсутствует при этом количество цитохрома Р-450 в печени морской свинки в 2-3 раза меньше чем в печени у крыс и соответственно меньше мощность ОСФ. Поэтому метаболические превращения ГК идут в несколько раз менее интенсивно, чем у крысы, и соответственно, других эу- и ксинобиотиков. Поэтому образование АФК и активных радикалов метаболизируемых веществ в печени и других тканях, имеющих системы детоксикации ОСФ, в несколько раз меньше, чем у крысы (Нечипоренко С.И. 1981).
Возможно именно эти обстоятельства определили другой спектр метаболитов-адаптогенов, которые образуются в печени морской свинки при стресс-воздействиях (вследствие низкой активности декарбоксилаз, стимулируемых АФК) – это глутамат, пролин, орнитин, глицин, цистеин и образуемый из этих аминокислот глутатион, который в условиях стресс-воздействиях интенсивно окисляется до ГSSГ и никуда из печени не транспортируется (Павлов В.А. 2000).
Кроме окисления глутатион интенсивно коньюгируется с метаболитами обезвреживания ГК, КХА, ПАУ и других эу- и ксенобиотиков с удалением этих веществ из клеток и тканей организма морской свинки.
У крыс мощная грубоволокнистая СТ, принимающая активное участие в процессах детоксикации – адсорбирует на своих волокнах токсичные вещества. Клетки СТ крыс способны производить большое количество УДФГК и ФАФС-веществ, коньюгирующих токсичные вещества и выводящие их из организма. Но не все токсичные вещества подвергаются такой коньюгации. Так, производные обмена ГК и ПАУ подвергаются гораздо более эффективной коньюгации и выведенной из организма с глутатионом, как это имеет место у морских свинок. У последних СТ нежноволокнистая синтезировать АК не способна, но при стресс-воздействиях она способна мобилизировать большие количества глутаминовой кислоты, глицина, пролина, необходимых для адаптивных перестроек в печени, прежде всего для биосинтеза глутатиона (Нечипоренко С.И. 1981, Голиков П.П. 1986, Павлов В.А. 2000).
Неспособность синтезировать АК приводит к тому, что при стресс-воздействии усиливается обмен углеводов не в сторону образования АК и УДФГК, а в сторону образования рибозы и включения ее в ГМФШ.
Смысл этих превращений состоит в том, что образующихся в ГМФШ НАДФН2 отдает свои восстановленности НАДН2, которые окисляются в ЦТК и дыхательной цепи ( Лабори А. 1970 )
Такой ход адаптивных метаболических перестроек без усугубления ОВП и образования АТФ возможен только при интенсивном функционировании ЦТК и дыхательной цепи, то есть усиления метаболических окислительных процессов углеводов по пути превращений в цикле Эмбдена-Меергофа-Кребса с интенсификацией работы ДЦ.
В тоже время у крыс в условиях стресса окисление углеводов по этому пути блокируется и направляется в активированный ГМФШ, обеспечивающий интенсификацию процессов окисления эу- и ксенобиотиков в ОСФ и образование повышенных количеств АФК для усиления свободнорадикального окисления в тканях свободных жирных кислот(СЖК), повышенного биосинтеза АК и метаболитов, используемых в биосинтезе соединительной ткани.
Следовательно у морских свинок усиленное образование глутатиона, его использование в антиоксидантных и детоксикационных механизмах наряду с усилением метаболизирования углеводов по пути Э-М-К и интенсификацией работы ДЦ – являются мощными антимутагенными факторами, что, видимо, и делает морскую свинку устойчивой к мута- и канцерогенезу, а также к хроническому эмоциональному стрессу. По своим механизмам ЭС протекает с усиленным образованием КХА и в гораздо меньшей степени ( хотя и постоянно ) ГК (т.е. ПАУ) и в процессе их метаболизирования образуются повышенные количества АФК в тканях и токсичные, повреждающие генетический аппарат метаболиты, что напоминает мута- и канцерогенные воздействия ПАУ (Середенин С.Б. 1980, Панин Л.Е. 1983, Моисеенко В.С., Загоруйко Л.И., Тодор И.П. 1985, Шабад Л.М. 1970, Шапот В.С. 1975, Худолей В.В. 1993, 1996).
Нам неизвестно по каким причинам морская свинка (в отличие от других видов млекопитающих) избрала этот путь адаптивных перестроек, очевидно, во многом это связано с особенностями обитания ее на островах недалеко от Южной Америки, но у приматов и человека, также не способных синтезировать АК и ориентирующихся в детоксикационных механизмах на повышенное образование глутатиона (его окисление и включение в процессы коньюгации) такая стратегия адаптивных перестроек, очевидно, связана с повышением устойчивости к ЭС, мута- и канцерогенезу. Именно эти обстоятельства, видимо, и определяют тот факт, что на протяжении десятков тысяч лет человек генетически несущественно меняется в отличии, например, от грызунов, собак, копытных и других животных (Шабад Л.М. 1970, Шапот В.С. 1975, Худолей В.В. 1993, 1996).
Имеются сведения, из которых следует, что интенсивность образования таурина в печени снижается в следующем ряду животных: крыса – собака – корова – лошадь – кошка – морская свинка – человек (Rossi 1987 и др.), то есть обратно пропорциональна устойчивости к ЭС мута- и канцерогенным воздействиям.
Как выше отмечалось, ткани морской свинки (печень, легкое, селезенка, мозг) содержат железа и серы в 2-3 раза больше, чем соответствующие ткани крысы. Повышенное содержание железа и серы обнаружено в мозге, печени приматов и человека. Железо и сера сосредоточены в основном в митохондриях аэробных тканей (Хьюз М. 1983) в виде железа – серных белков и цитохромов ДЦ. Высокое их количество говорит об ориентации метаболизма на путь Э-М-К с высокой активностью ДЦ и в условиях стресс-воздействий об активизации, а не угнетения этих систем.
Наиболее аэрируемой тканью организма является мозг, содержащий наибольшее количество митохондрий и железо-серных комплексов по сравнению с другими тканями организма (Уайт А. 1981). Считается, что обмен нервной и мозговой ткани в принципе одинаков у всех животных (Кричевская А. А, 1982). Однако в условиях стресс-воздействий у различных видов животных аэрация мозга различна, как различны и субстраты для энергообразования (Панин Л.Е. 1983).
Так, мозг крысы в условиях стресс-воздействия вполне может окислять СЖК с образованием большого количества АТФ, и достаточно устойчив к глубокой гипоксии. В тоже время мозг человека, приматов и морской свинки в условиях стресс-воздействия облигатно зависим от поступления углеводов и достаточной степени аэрации, особенно при ЭС.
При этом необходимо указать, что внутри каждого вида имеются особи существенно отличающиеся друг от друга по чувствительности к различным стресс-воздействиям.
Из имеющихся литературных данных (Панин Л.Е. 1983) известно, что животные, также как и человек могут относится к разным защитно-адаптивным типам. Так, выделяют высоко устойчивых особей (как между видами так и внутри каждого вида) к острым и интенсивным стресс-воздействиям, которые гораздо менее устойчивы к воздействиям менее интенсивным, но длительным. Это так называемые «спринтеры». Другой крайний тип – это особи устойчивые к длительным и мало интенсивным воздействиям (особенно к эмоциональному стрессу), но менее устойчивые к острым и интенсивным стресс воздействиям (особенно физическим повреждающим факторам – травма, температура, токсические воздействия и т.д) — “стайеры”. Между этими типами существуют различные промежуточные типы. (Казначеев В.П.1984, Семагин В.Н1988., Робу А.И. 1989, Яковлев Г.М.1990, Судаков К.В. 1984,1997).
Нам удалось установить (Павлов В.А. 2000), что для «спринтеров» более характерен N-тип защитно-адаптивных механизмов (печеночный соматический метаболический тип с преобладанием в адаптивных механизмах метаболитов богатых азотом), ориентирующийся в основном на перестройку метаболических процессов в печени с выделением в кровь таких веществ как таурин, аргинин, полиамины, мочевина, то есть веществ, содержащих в повышенных количествах азот. Из метаболитов, содержащих серу при таком защитно-адаптивном типе, преобладает образование таурина, в котором сера окислена до SO 3.
Для «стайеров» более характерно наличие S-типа с большим значением защитных механизмов перераспределения аминокислот из соединительной и нервной ткани. Здесь преобладают такие метаболиты как глутаминовая кислота, пролин, глицин, цистеин и глутатион, то есть вещества, содержащие в повышенном количестве серу, в которых она находится в восстановленном состоянии: SH.
Выше уже отмечалось, что стресс-воздействия влияют на защитно-адаптивные системы организма, стимулируя их. Соответственно повышается устойчивость к повреждающим воздействиям (эустресс). И напротив, когда происходит истощение защитно-адаптивных систем, развивается патологическое состояние (дистресс). Частые, превышающие адаптивные ресурсы, повреждающие воздействия могут привести к гибели организма.
Организмы с преобладанием N-типа (“крысы”) более устойчивы к острым и интенсивным стресс-воздействиям (перепады температур, физическая нагрузка, возбудители инфекционных заболеваний и т.д.). Это связано с тем, что вещества-адаптогены, образующиеся в повышенном количестве при таком защитно-адаптивном типе обладают мембраностабилизирующим эффектом и рядом других особенностей, необходимых при грубых и интенсивных стресс-воздействиях. То есть этот защитно-адаптивный тип направлен на “противоудар” — на сопротивление.
Организмы с преобладанием S-типа защитно-адаптивных систем малоустойчивы к интенсивным и острым, но гораздо более устойчивы к малоинтенсивным и длительным стресс-воздействиям. Малоинтенсивный но хронический стресс требует соответствующей ему стратегии защиты. Важнейшее значение имеет то обстоятельство, что такие стресс-воздействия сопряжены с постоянным повышением в тканях активных форм кислорода и свободных радикалов (хотя и в гораздо меньших количествах, чем при острых и интенсивных стресс воздействиях), которые оказывают повреждающее воздействие на мембраны клеток, особенно митохондрий, ответственных за образование АТФ и обеспечение организма энергией. Для защиты клеток в такой ситуации необходимы вещества, образующиеся в большом количестве в мозге и нервной ткани. Из этих веществ-адаптогенов образуется глутатион, который и является главным фактором защиты при хроническом стресс-воздействии. Экспериментально доказано, что наибольшее отрицательное влияние оказывает хронический эмоциональный стресс, при котором повреждаются как митохондрии, так и ядерный аппарат клетки (Панин Л.Е. 1983). Это приводит к повышению количества мутаций и канцерогенным воздействиям, что сопровождается развитием различных патологических состояний, вплоть до возникновения злокачественных опухолей (Худолей В.В. 1996). Главным защитным механизмом в данной ситуации является глутатион. На этой основе формируется другой тип защитно-адаптивных систем — тип “избегания”, когда защитно-адаптивные системы направлены не на сопротивление повреждающему воздействию, а на уход от него. Главенствующее значение в такой стратегии имеет развитый мозг и другие отделы нервной системы.
Таким образом, развитие высокоорганизованной нервной системы, мозга, является средством приспособления организма к хроническим стресс-воздействиям, прежде всего к эмоциональному стрессу, поскольку именно в нервной системе и образуются вещества (глутаминовая кислота, цистеин, глицин), необходимые для образования глутатиона. Эмоциональный стресс в силу этой причины является мощным фактором эволюционного развития, так как является фактором стимуляции цефализации, существенно ускорившей эволюцию. Так, в определенной мере, эволюционное восхождение в плане совершенствования адаптивно-защитных механизмов среди млекопитающих можно проследить от грызунов (крысы, мыши) и до приматов. Известно, что крыса является своеобразным высоко адаптивным биологическим реликтом (Ж. Сент – Илер 1968), хорошо переносящим острые и интенсивные повреждающие воздействия (температура, токсические вещества, радиация и т. д…). Важнейшее значение в защитных механизмах к повреждающим воздействиям у крысы имеет печень, где в большом количестве образуются адаптогены (аргинин, полиамины, мочевина и ряд других), которые накапливаются в важнейших защитно-адаптивных системах организма (кроветворная, иммунная, детоксикационная и др), но особенно в больших количествах в тканях у крыс обнаруживается таурин -50% и более от всех аминокислот. Промежуточное положение занимают другие млекопитающие. Так, например, если у крыс содержание таурина максимальное и максимальная устойчивость к острым и интенсивным стресс-воздействиям, то, для сравнения , у собак содержание таурина гораздо меньше, и меньшая устойчивость к аналогичным стресс-воздействиям. Еще меньшая концентрация таурина у кошек. Одно из последних мест в этом ряду(по количеству таурина в тканях) занимают приматы и человек. Но у человека имеются два полюса защитно-адаптивных систем-N-тип и S-тип, что делает его гораздо более адаптивным по сравнению со всеми живыми существами по отношению к воздействиям факторов окружающей среды.
Таким образом, завершает эту пирамиду человек, имеющий максимальные защитно-адаптивные возможности. Эти возможности состоят из двух полюсов — “печеночного” (N-тип) и мощного “мозгового”(S-тип), что позволяет иметь среди человеческой популяции широкий спектр различных вариантов защитно-адаптивных механизмов. В плане экспериментальных исследований защитно-адаптивных типов большую ценность имеют опыты, проведенные нами на крысах и морских свинках. У этих животных защитно-адаптивные типы являются как бы крайними вариантами, присущими для человека.
Про крыс мы уже упоминали – это крайний печеночный N-тип. Другим крайним типом является морская свинка. По своим обменным процессам, в силу некоторых обстоятельств, похожа на приматов и человека, что могло быть обусловлено особенностями ее жизни на островах, в определенных условиях. Морская свинка имеет ярко выраженный S-тип (перераспределительно-мозговой), но мозг у нее очень маленький, а “печеночный” N-тип выражен очень слабо. Для морской свинки характерна повышенная чувствительность к различным острым и интенсивным стресс-воздействиям (радиации, перепадам температур, интоксикациям, заражению многими возбудителями инфекционных заболеваний и т.д.). Но она имеет высокую устойчивость к факторам, повреждающим ее генетический аппарат (хронические интоксикации, мутагенные и канцерогенные воздействия).
Важнейшими факторами, определяющими устойчивость, как морской свинки, так и человека к хроническим малоинтенсивным стресс-воздействиям, эмоциональному стрессу, мутагенным и канцерогенным воздействиям являются образующиеся в их организме, в большом количестве, вещества, защищающие генетический аппарат от мутагенных (вызывающих мутации) и канцерогенных (вызывающих опухоли) веществ. К таким защитным метаболитам относятся глутатион, цистеин, и ряд других, серосодержащих аминокислот и их производных.
Такие метаболиты или их предшественники, в большом количестве, образуются в мозге и нервной системе. А также они содержатся в соединительной ткани, из которой при стрессовых воздействиях могут перераспределяться в защитно-адаптивные системы организма (перераспределительный тип).
Чем более высоко развит вид в плане совершенствования его ЦНС и мозга, тем более различна реакция его представителей на то или иное стресс-воздействие. Так, мы уже отмечали, что исследователи выделяют «спринтеров» и «стайеров» по устойчивости к острому и интенсивному и малоинтенсивному, но хроническому стрессу (Казначеев В.П., 1984, Юматов Е.А. 1995, Чирков А.М. 1989).
Однако необходимо констатировать, что в принципе все млекопитающие, находящиеся в диких условиях природы, с каждым годом все более и более теряют свою адаптивность и исчезают как виды (Новоженов Ю.И. 2005).
Это касается также и многих других представителей флоры и фауны. Все это происходит оследствие колоссальной антропогенной нагрузки и чрезвычайно высокой адаптивности как вида человека. Но по мере нарастания техногенной нагрузки на биосферу и загрязнения окружающей среды выбросами различных производств, транспорта, отходами производства и быта людей они и сами начинают терять адаптивность. Главный показатель падения адаптивности – резкое падение рождаемости, особенно технически высокоразвитых народов (Панарин С.А. 1998, П. Бьюкенен 2003, Новоженов Ю.И. 2005).
Здесь необходимо остановиться на том, что человек, имеющий наиболее развитую ЦНС и мозг среди всех других представителей живой природы внутри своего вида имеет максимальные различия по отношению к воздействию различных стресс-факторов (Панин Л.Е. 1983, Казначеев В.П.).
Так, из многочисленных научных исследований да из наблюдений в быту хорошо известно, что отдельные люди хорошо переносят острые и интенсивные стресс-воздействия, то есть являются «спринтерами», а другие – малоинтенсивные, но длительные — хронические – «стайеры». Так, среди «спринтеров» можно отметить людей, хорошо переносящих физическую нагрузку, они, как правило, хорошо концентрированы, имеют мощные тормозные процессы на уровне ЦНС – хорошо засыпают, уравновешены, внимательны (Казначеев В.П., 1984, Юматов Е.А. 1995, Чирков А.М. 1989).
Такие люди чаще устойчивы к интенсивной физической нагрузке, холоду, инфекционным заболеваниям, воздействию токсичных веществ. Более того, они склонны к употреблению алкоголя и курению. Но они чаще всего не склонны к интенсивной умственной деятельности, плохо переносят хронический эмоциональный стресс – следствием чего является алкоголизм, психические расстройства.
Другой крайний тип «стайеры» – это люди чаще умственного труда, плохо переносящие острые и интенсивные физические стресс-нагрузки, имеют слабые тормозные процессы на уровне ЦНС, не склонны к употреблению алкоголя и курению. Однако такой тип ЗАМ устойчив к хроническому эмоциональному стрессу, высоким умственно-коммуникативным нагрузкам (Панин Л.Е. 1983, Семагин В.Н. 1988, Менделевич В.Д. 2002 Судаков К.Ф. 1997).
Согласно данным токсикологических и патофизиологических исследований повышенную устойчивость к различным интоксикациям и повреждениям имеют люди с мощной системой ОСФ и повышенным количеством Р-450 в печени, функция которых обеспечивается мощным ГМФШ в печени (Голиков С.И 1986).
Таким образом, люди, являющиеся в большей степени «спринтерами»ориентируются в своих защитно-адаптивных механизмах (ЗАМ) на доминирующую роли печени, тогда как у «стайеров» в большей мере доминирует мозг и в целом надо констатировать, что у человека в отличии от животных имеются как бы два полюса образования метаболитов-адаптагенов – печень и мозг. Это определяет широкий спектр защитно-адаптивных типов (ЗАТ) в человеческой популяции и делает ее гораздо более адаптивной по сравнению с популяциями и тем более отдельными особями всех других видов.
Воздействие на организм вирулентной микрофлоры при развитии и прогрессировании инфекционного процесса (например, мышиный тиф, туберкулез, трипаносомоз и др.) снижает уровень цитохрома Р-450 в печени и наоборот относительно легко перенесенная инфекция и выздоровление повышают уровень и соответственного детоксикационные ресурсы организма (Сокирко Т.А. 1990, Малюк В.И. 1984, Bartatoli G.M. 1997). В печени образуются белки системы комплемента, существенно повышающие антимикробные свойства иммунной системы («Иммунология» Петров Р.И. 1993).
Точно также и воздействие мута-канцерогенных ксенобиотиков типа ПАУ в небольших субтоксичных для данного организма дозах повышает иммунные и детоксикационные ресурсы (Ковалев И.Е. 1977, 1984). То есть людей такого типа условно можно назвать «печеночники».
И, напротив, в условиях хронического эмоционального стресса главенствующие значения в ЗАМ имеют не детоксикационно-метаболические ресурсы печени, которые включают в себя системы ОСФ – цитохром – Р-450 с интенсивным биосинтезом таких веществ как таурин и стабилизаторы мембран — полиамины, а такие адаптогены как глутамат, ГАМК, ГОМК, карнозин – глутамил пептиды (например Г-SH), аргинин, мочевина, защищающие, например, миокард от повреждающего воздействия (Меерзон Ф.З. 1984, 1997, 1999). Все эти вещества образуются в головном мозге в повышенных количествах и в условиях стресс-воздействия могут защищать как саму ткань мозга так и перераспределяться в другие органы (для чего есть специальные транспортные системы). Оказывают защитный эффект, например в сердечно-сосудистой системе, миокарде. (Раевский К.С., 1986, Меерзон Ф.З. 1999).
То есть, люди с преобладанием такого защитно-адаптивного типа механизмов являются «мозговиками».
Однако деление это условно, так как мозг и печень — два наиболее крупных паренхиматозных органа, которые составляют примерно по 2% от массы тела человека, дополняют и координируют функцию друг друга в условиях адаптивного ответа на стресс-воздействие. Так, печень обеспечивает мозг серосодержащими метаболитами-аминокислотами, для которых в ГЭБ существуют особые транспортные системы. Таурин и цистеин активно поглощаются мозговой тканью. Таурин обладает мощным общим тормозным действием, координирующим многие медиаторно-регуляторные эффекты в коре и подкорковых структурах. Мозговая ткань содержит большие количества серосодержащих аминокислот – таурина, цистеина, цистатионина, изетеиновой кислоты (Раевский К.С. 1986, Нефедов Л.И. 2001).
В свою очередь в головном мозге образуется большое количество – азотсодержащих метаболитов – адаптогенов широкого спектра – глутамата и его производных ГАМК, глутамин, мочевина, N-ацетилглутамат, глутамин транспортирующихся в печень и стимулирующих здесь адаптивные процессы, например, биосинтез мочевины стимулируемый N-ацетилглутаматом, биосинтез нуклеиновых кислот и полиаминов, стимулируемых N-ацетиласпартамом и глутамином, поступающих из мозга и т.д. (Кричевская А.А. 1982, Линдауэр Р. 2003, Раевский К.С. 1986, Яковлев Г.М. 1990).
Однако, несомненно, и то, что в адаптивных процессах в зависимости от условий среды, в ответ на стресс-воздействие может преобладать либо мозг, либо печень и соответственно люди с таким преобладанием могут условно обозначаться – «мозговики» или «печеночники».
В проведенных нами исследованиях спектра аминокислот-адаптогенов у 5% людей он напоминает таковой у морской свинки. У 42% он напоминает таковой у крысы. 53% различные промежуточные типы.
В первом случае это были чаще всего люди умственного труда, с относительно слабым здоровьем. Если они заражались туберкулезом, то он протекал у них тяжело, а лечение противотуберкулезными препаратами часто давало побочные эффекты в виде гепатитов, поражений нервной системы, слухового аппарата и т.д.
Во втором случае – это чаще всего люди физического труда, не склонные к напряженной умственной и творческой деятельности. У людей второго типа туберкулез развивался либо после длительного пребывания в заключении или воздействия других сильных и длительно действующих стрессовых факторов.
Соотношение аминокислот адаптогенов и некоторых их производных, которые определяются в крови здоровых и больных различными заболеваниями людей, дает определенные коэффициенты. По этим коэффициентам можно рассчитать принадлежность индивидуума к тому или иному адаптивному типу и метаболический адаптивный ресурс организма. Учет полученных результатов позволил назначить и провести целенаправленную коррекцию метаболических и адаптивных нарушений и существенно повысить эффективность лечения, например, тяжелых форм туберкулеза. (Павлов В.А. 2000).
Полученные результаты позволяют говорить о том, что у людей, спектр аминокислот-адаптогенов которых в определенной степени соответствует таковому у крысы, эти вещества нарабатываются в основном в печени. У людей спектр аминокислот-адаптогенов в крови напоминающих таковой у морской свинки эти метаболиты образуются в основном в мозге, поэтому первых условно можно обозначить как «печеночники», а вторых как «мозговики». Люди 2-го типа – чаще всего люди умственного труда, чувствительные к инфицированию МБТ, люди 1-го типа – чаще всего люди физического труда, заболевают туберкулезом лишь после длительных неблагоприятных условий жизни – длительный стресс, длительные сроки заключения (Павлов В.А. 2000).
Весьма интересны сведения о становлении человека как обладателя высокоадаптивного типа защитных механизмов.
Так, известным русским психологом, физиологом, антропологом и историком Поршневым Б.Ф. была выдвинута достаточно аргументированная концепция о том, что промежуточный этап перехода от обезьяны к человеку имел те особенности, что существенно менялись форма и объем мозга. И если у наших предков, еще целиком находящихся в царстве животных, размеры мозга (понгиды) соответствуют примерно современным шимпанзе, то уже следующий этап перехода – троглодиты размеры мозга значительно увеличиваются доходя до максимальных размеров у палеоантропов и неоантропов, что можно видеть из приведенной таблицы №1, взятой из монографии Поршнева Б.Ф. (по Кочетковой В.И.).
Табл. №1 Размеры мозга по эндокранам ископаемых черепов:
| Понгиды | Троглодиты | Гоминиды | ||||
| в мм. | шимпанзе | австралопитек | археантроп | палеоантроп | неоантроп | совр. человек |
| длина мозга | 106 | 125 | 155 | 174 | 186 | 167 |
| лобная доля | 81 | 92 | 110 | 128 | 135 | 129 |
| теменная доля | 71 | — | 108 | 118 | 116 | 124 |
Показатели, составленные по эндокранам (вдавлениям) ископаемых черепов имеют самый большой размер у неоантропов (кроманьонцев) – ископаемых гоминид. И он уже достаточно большой у предшествовавших им троглодитов – палеоантропов (неандертальцев), еще собственно к людям (гоминидам) не относящихся. Но, как видим, и у тех и у других меньшие размеры теменной доли, в которой сосредоточены речевые центры.
Удивительно мощное адаптивное значение мозга (наработка метаболитов-адаптогенов) его значительное увеличение у неандертальцев и особенно кроманьонцев имело ведущее значение в совершенствовании механизмов адаптации.
Как утверждает Поршнев Б.Ф. троглодиты питались падалью (крупных животных, белковая пища, способствующая развитию мозга) и имели большой и мощный мозг, который в функциональном плане выполнял прежде всего функцию суггестии (внушения, психологического воздействия) на хищных животных и себе подобных для защиты от их нападения в виду того, что предки человека естественных средств защиты и нападения (клыки, когти, рога) не имели.
Именно поэтому человек (и его предки) никогда не были жертвами и объектами охоты для хищников. Последние, как правило, боятся человека, не выносят его взгляда. И видимо, боялись и еще более не выносили взгляда и выкриков типа кыш, фу, брысь! (его предков – мозговиков-суггесторов).
Возможно, что эти предки благодаря своему супермозгу лучше, чем мы чувствовали гармонию мира и жили непосредственно в естестве природы в гармонии с ней.
Но ситуация постоянно менялась, менялся окружающий мир, его физико-химические параметры, прежде своего снижение температуры (наступление ледникового периода) и изменения других параметров биоциноза. И преобладание чисто мозговой адаптивности стало явно недостаточно. Одним из таких факторов можно отметить то, что предки человека научились использовать огонь, поэтому на них резко возросло воздействие продуктов сгорания и перегонки органического материала. В этих условиях необходимым фактором адаптивности стала печень, обладающая мощным детоксикационным потенциалом и нарабатывающая мощные защитные адаптогены, способствуя тем самым реализации адекватных повреждающим воздействиям адаптивных перестроек.
Как мы отмечаем выше самым мощным адаптагеном печени является таурин, обладающий общим тормозным воздействием на возбудимые ткани.
Как установлено в наших исследованиях в условиях стресс-воздействия таурин стимулирует и регулирует функцию кроветворной, иммунной, антиоксидантной, детоксикационной систем подопытных животных (Павлов В.А 1996, 1999, 2000). Не менее значимо его воздействие на функцию и состояние метаболических систем, НЭС и мозга. Содержание таурина в мозге, щитовидной железе, надпочечниках в условиях стресс-воздействия возростает в 2-3 раза. Обладая мощным тормозным воздействием за счет стабилизирующего влияния на электролитный обмен мембраны а также за счет антиоксидантной ее защиты таурин повышает ее окислительно-восстановительный потенциал и снижает вобудимость. Таурин является важным фактором усиления тормозных процессов в мозге.
Здесь необходимо отметить, что (согласно данным Поршнева Б.Ф.) само по себе понятие человеческого общества (общины, рода и т.д.) стало возможным только с возникновением речи и соответственно социальной психологии. Появилось такое понятие как «мы» и «они» – чужие и на первых порах они – чужие как раз и выступали сохранившиеся еще предшественники людей – неандертальцы или ранние кроманьонцы – которые не знали еще общины, были распространены диффузно в природных местах обитания (имеющих достаточное количество белковой пищи – падали и более-менее пригодные климато-географические условия).
Формирование межнейронного взаимодействия в процессе адаптивных перестроек в мозге во многом сопряжено с усилением тормозных процессов (Поршнев Б.Ф.). Поэтому формирование «мыслительного ядра» или «доминанты Ухтомского», определяющих мыслительно-мировоззренческие особенности человека и лежащие в основе формирования эмоций, особенностей мышления и, в конечном счете, личности, по современным представлениям, определяется такими медиторами ЦНС и мозга как глутамат, ГАМК, ГОМК, катехоламины, дофамин, серотонин, эндорфины, энкефалины, таурин и ряд других веществ, обладающих помимо медиаторной также функцией адаптогенов (Вальдман А.В. 1976, 1979, Бехтерева Н.П. 1985, Судаков 1997, 2004, Менделевич В.Д. 2002, Линдауэр Р. 2003, Раевский 1987, Меерзон Ф.З. 1984, 1987, 2004 и др.). Образование этих веществ может происходить как в ЦНС и мозге так и в печени – основных центрах наработки адаптогенов в организме. И преобладание того или другого центра будет влиять на формирование важнейших защитно-адаптивных типов человека и окружающего его общества.
Здесь необходимо отметить, что по данным Раевского К.С. 1986, Поршнева Б.Ф. 1986, Нефедова Л.И. 1992, 1999, Короедова Л.М. 1994, 1998 у больных с нарушениями вследствие заболеваний и травм в лобной и теменной долях, то есть наиболее молодых структур мозга, развиваются психопатические состояния со значительным снижением адаптивности и снижением концентрации таурина в ткани головного мозга и крови. Введение таурина оказывает нормализующее воздействие.
Согласно фундаментальным разработкам отечественной школы физиологов Павлов И.П., Анохин П.К., Ухтомский А.А., Веденский И.Е. именно тормозные процессы лежат в основе формирования 2-ой сигнальной системы и формирования речи как таковой.
Видимо усиление роли печени в адаптивных механизмах способствовали развитию и становлению человека современного вида. Процентное соотношение мозг-печень неандертальца и кроманьонца 3% / 2% у современного человека 2% / 2% (у животных 1% / 4-6%). Таким образом наработка печенью тормозных адаптогенов, видимо, способствовала формированию лучшей адаптированности к изменяющимся условиям среды, повышению координированности движений и соответственно росту теменной и лобной долей мозга, что в дальнейшем сделало возможным формирование речи, центры которой находятся рядом с двигательными центрами в лобной и теменной долях. С возникновением этого наиболее совершенного канала коммуникации начинает формироваться человеческое общество, общественная психология, да и соответственно, сам человек в современном смысле. Речь, воздействуя на соответствующие мозговые структуры, через ретикулярную фармацию и другие отделы подкорковых структур мозга воздействует на весь организм, а в свою очередь органы организма, прежде всего активно участвующие в механизмах адаптации (печень, кроветворения, иммунная система и др.) в свою очередь действуют на мозг (Поршнев Б.Ф. 1975, Судаков К.В. 1997).
Поэтому человек, в принципе, выходит уже на другой уровень адаптивных механизмов – коммуникативно-метаболический. В котором на равных правах участвуют как вербально-коммуникативное поле, так и адаптивно-метаболические процессы важнейших адаптивных систем организма. То есть, состояние социальной психологии так и адаптивно-метаболические ресурсы членов того или иного общества или популяции взаимосвязаны.
Появившиеся первые люди – общинники, бывшие уже в большей степени печеночниками с более совершенными тормозно-адаптивными процессами и лучшими адаптивными качествами по отношению к меняющимся климато-географическим условиям, уступали предшествовавшим им формам (неандертальцы, ранние кроманьонцы) лишь в том, что они уже утрачивали возможности к суггестии. И менее совершенные адаптивно-физически сохранившиеся неандертальцы при встрече с людьми, являясь мощными суггесторами, действовали на них как удав на кролика, а возможно и вообще употребляли их в пищу, что, возможно, и лежало в основе каннибализма.
Поршнев Б.Ф. считает, что именно эти обстоятельства и послужили причиной многотысячекилометровых миграций появившихся людей общинников, бегства их от малоадаптивных но высокосуггестивных предшественников. И действительно, каких-то других разумных причин для того, чтобы преодолевать с риском для жизни многотысячекилометровые расстояния по суше (пересеченной местности) и океану, кроме как объяснить смертельным ужасом перед своими предшественниками представить невозможно, других причин до сих пор не найдено.
Расселившись на большие пространства люди-общинники (печеночники) довольно легко адаптировались к местным климато-географическим условиям всех континентов (кроме Антарктиды) тогда как предшествовавшее им формы мозговиков-суггесторов бесследно исчезли ввиду их малой адаптивности.
С этого момента начинается новый этап развития живой материи – собственно уже человеческая история, ибо уже животный мир (исключая прокариот) конкурировать с ними в плане совершенствования защитно-адаптивных механизмов уже никак не мог.
Исторический анализ развития человеческого общества говорит о том, что процесс этот начинается с разложения его первичной матрицы- общины с появлением социальной надстройки и становления государств различного типа в зависимости от исторически-цивилизационного среза. Однако закономерность всех его этапов- возникновения, становления, достижения своего максимального развития, упадка и исчезновения во многом одинаковы, как говорил арабский мыслитель Ибн-Хальдун: на развалинах старых империй новые варвары создают новые государства и империи. В основе процессов общественного развития лежит энергия адаптивности первичной матрицы того или иного народа – общины. На пути исторического развития защитно-адаптивным механизмам этого народа предстоит трансформироваться от преобладания адаптивных механизмов к естеству климато-географических воздействий к преобладанию ЗАМ к стресс-воздействиям общественно-коммуникативным развитого социально-коммуникативного общества. Это уже совсем другие стресс-воздействия в результате которых возникают совсем другие люди.
Находясь в условиях естества первичной крестьянской или кочевнической общины человек связан многими тормозными ограничениями общины – табу, он целиком зависит от общины и не может реализоваться как личность – главные его качества – сила, выносливость, терпимость к решениям общины, то есть он высокоадаптивный к условиям естественного существования «печеночник». Это подтверждает и высокая рождаемость (плодовитость) у народов находящихся на этой ступени развития их высокая устойчивость к инфекциям, различным трудностям и лишениям.
У человека, ступившего на социально- исторический путь развития, преобладают уже совсем другие адаптивные механизмы – прежде всего устойчивость к социально-коммуникативному стрессу.
Изменение спектра стресс-воздействий меняет коммуникативно-метаболические адаптивные системы как общества так и отдельных индивидуумов, они становятся все более раскрепощенными (свободными). На метаболическом уровне это связано с постепенным снижением роли печеночных защитно-тормозных( таурин, полиамины, мочевина) адаптагенов и возрастанием – мозговых, определяющих большую мобильность и лабильность (переключаемость) мембрано-функциональных структур мозга и других органов – то есть происходит трансформация от «печеночников» к «мозговикам» – более адекватным эмоционально-коммуникативным и дугим. антропогенным стресс-нагрузкам.
Однако многолетний и тем более многовековой социально-исторический «стаж» приводит человека и в целом общественный организм к потере адаптивности – резко падает рождаемость, возрастает число психических и других заболеваний, социальная психология ориентируется на потребительство, гедонизм, различные извращения, в том числе гомосексуализм (особенно элитарные слои общества), процветают алкоголизм, наркомания, и т.д. В результате бывший великий народ деградирует, на смену ему должен придти другой более здоровый народ и основать новое государство, располагающее соответствующим адаптивным ресурсом развития.
Таким образом, структурно-функциональное и метаболическое совершенствование нервной системы, и прежде всего мозга, привело к формированию важнейшего инструмента адаптации во многом определяющего резкое ускорение эволюционных процессов. Этот факт был отмечен еще в середине 19 века, американским ученым Д.Д. Дана, который сформулировал концепцию о цефализации, то есть развития центральной нервной системы, и прежде всего мозга, как основы эволюции высокоорганизованных организмов. Причем обратного хода как цефализация, так и эволюция не имеют.
Концепция о цефализации предполагает значительную стимуляцию перераспределительно-мозгового типа как фактора значительного повышения адаптивных возможностей организма к эмоциональному стрессу и другим антропогенным стресс-воздействиям также как и популяции в целом, путем интегрирования ее защитно-адаптивных механизмов в виде культуры, как формы адаптации. В человеческом сообществе это выражается в возникновении все более совершенных форм культуры как фактора адаптации популяции, общества и цивилизации в целом.
В книге Новоженова Ю.И. “Филитическая эволюция человека” убедительно обосновано, что человеческая популяция, общество и цивилизация имеют свои определенные уровни адаптивности и соответственно им реагируют на стресс-воздействия окружающей среды. То есть, эти уровни организации живой материи, так же, как и ее отдельные организмы, можно с полным правом считать своеобразными живыми системами. Однако их эволюционный процесс реализуется уже не в виде совершенствования морфологически-фенотипических признаков, а виде возникновения и совершенствования культуры, как основной формы адаптации цивилизованного человеческого общества. И более адаптивные в этом плане популяции занимают лидирующее положение в развитии цивилизационного процесса. Наши исследования подтверждают эти положения и развивают их.
Суммируя изложенное можно отметить, что в человеческом обществе среди стресс-воздействий наиболее мощное стимулирующее значение на процессы цефализации имеет эмоциональный стресс, который имеет определенное сходство с мутагенными и канцерогенными воздействиями. Но эволюционные процессы происходят уже не только и не столько на уровне функционально-метаболического, генетического и фенотипического совершенствования, а на уровне стимуляции цефализации и развития более совершенной культуры, более гармоничными общественными отношениями, то есть, с защитно-адаптивными метаболическими механизмами, связанными в большей мере с совершенствованием перераспределительно-мозгового типа адаптации. Но популяции различных человеческих сообществ, так же, как и люди в них, бывают разные.
Одни хорошо адаптируются к острым и интенсивным стресс-воздействиям, они сильны и выносливы (“спринтеры”). Другие гораздо менее выносливы и устойчивы к различным интенсивным, повреждающим факторам. Но они умны и могут быть творчески одарены (“стайеры”).
Эти подходы позволили нам понять, что у людей, так же, как и у животных существуют два крайних защитно-адаптивных варианта: тип сопротивления (тип “крысы”) и тип избегания (тип “морской свинки”), подобно тому, как еще Гиппократом были выделены различные типы темперамента (сангвиник, флегматик, меланхолик, холерик). Сопоставляя полученные нами в эксперименте результаты с защитно-адаптивными механизмами у различных млекопитающих животных и у человека, мы пришли к выводу, что все возможные варианты защитно-адаптивных механизмов находятся между двумя этими крайними типами, в той или иной мере приближаясь к ним.
Полученные результаты дают основания разделить типы защитно-адаптивных возможностей организма на метаболически-печеночный и перераспределительно-мозговой, которые используют в своих антиоксидантных, детоксикационных процессах, а также в процессах стимуляции и регуляции генетического аппарата и метаболической сети различные метаболиты-адаптогены (схема 2). Применительно к человеку, такое различие будет определять разнообразие адаптивно-защитных механизмов, темперамента, функциональных и мыслительных способностей индивида. Люди с крайним “печеночным” типом (тип “крысы”) устойчивы к различным повреждающим факторам, склонны к употреблению алкоголя (лобилизатор мембран) и курению (защита ксенобиотиками своих собственных биологически активных регуляторов). Это тип сопротивления. Но такие люди не склонны к творческой и интенсивной мыслительной деятельности.
Другой тип — “мозговой” (тип “морской свинки”). У такого типа адаптогены образуются преимущественно в мозге и соединительной ткани, за счет чего развитие мозга усиливается, и соединительная ткань у такого типа рыхлая и слабая. Такой тип людей склонен к умственной деятельности. В то же время слабость ‘печеночных” механизмов защиты у “мозгового” типа определяет гораздо меньшую устойчивость к повреждающим факторам, и другим неблагоприятным воздействиям(это тип избегания).
2. ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ПСИХОЛОГИЧЕСКОЙ АДАПТИВНОСТИ
2.1 ВВЕДЕНИЕ К ЦЕРЕБРАЛЬНОЙ ЭВОЛЮЦИИ КАК ОСНОВЫ АНТРОПОГЕНЕЗА
Память о далеком прошлом человечества никогда не перестанет беспокоить людей. Выбирая на перепутье свою дорогу в будущее, невозможно не вспомнить пройденное. Даже отдельному человеку эта процедура не всегда представляется простой – все, что пережито часто оставляет в памяти эмоционально окрашенные следы. Приятно вспомнить то, что приятно вспомнить, но все ли было так хорошо в нашем прошлом, если механизм вытеснения (активного забывания) не позволяет с легкостью вспомнить всю так недавно прошедшую жизнь во всех её подробностях? Эти эмоции при пересмотре индивидуального прошлого захватывают человека, заставляют снова пережить крутые повороты судьбы. Невозможно без гнева и боли, радости и предвкушения счастья пережить то, что когда-то вызывало те же чувства. Непереносимая боль же забывается настолько, что события связанные с ней могут быть напрочь вычеркнуты из памяти. И тогда анализ пройденного пути будет нести в себе определенного рода ошибку. Будучи положена в основу рассуждений при планировании будущего, эта ошибка станет основой для неудач. Для отдельного человека неудача – если она не приводит к летальному исходу – повод для размышления, и, возможно, отправная точка для совершенствования.
Для человеческого общества ошибка в выборе направления развития означает на сегодняшний момент, может быть, и не гибель человечества, а заодно и современной природы, но, по меньшей мере, утрату смысла существования для абсолютного большинства ныне существующих людей. Те же, от кого реально зависит такой выбор, делают его, не ставя в известность остальных о предпосылках, лежащих в основе их планирования. Допустить, что богатое меньшинство знает, что делает, мне не позволяет мое знание. Этим знанием я и делюсь в настоящей работе.
Прежде всего, хотелось бы понять, что для человека может составить счастье. Речь идет не об отдельно взятом человеке, речь идет о строе общества, при котором большинство людей могли бы родиться и жить и умереть счастливыми. Возможно ли такое в настоящее время, или в будущем, ближайшем или отдаленном, мы можем предположительно сказать, только обратив взор в прошлое. У нас нет параллельного человечества, на опыте которого мы могли бы учиться. Человечество это мы, и другого нет. Только опыт нашего собственного прошлого может нам подсказать, что может получиться из наших попыток смоделировать будущее. Для этого, прежде всего, надо знать, что для человека является наиболее естественным. Вмешиваясь в ход событий, неплохо было бы иметь представление о том, каким был бы этот ход без нашего участия. Увидеть это не представляется возможным. Мы можем только погрузить взор в прошлое и отследить все попытки искусственного вмешательства в течение процессов протекающих в природе.
Человек возник в природе естественным путем. Химические, физические, биологические процессы протекали без вмешательства извне, и эти процессы привели к появлению на планете человека. Гипотезу сознательного творения я отвергаю, как излишнюю, ни бог, ни инопланетяне не кажутся мне необходимыми для антропогенеза.
В то же время, интенсивные исследования антропогенеза в 20 веке сохранили достаточно неясностей, для того, чтобы негодяи, заинтересованные в прогрессе мракобесия, и те, кто готов скинуть с себя ответственность за собственную безответственность на потусторонние силы могли спекулировать на недостатке научных знаний.
Известно, что человек современного вида появился на планете около 10 000 лет назад. Сегодня этот вид расселился по всей планете, от полюса до полюса. Есть места, где люди живут в природе, очень мало используя достижения человечества в области науки и техники. Рис.1 и 2.
Тасадеи весь день проводят вместе, а так как дел и развлечений у них немного, они значительную часть времени сидят молча тесными кучками, как, вероятно, сидели и первые люди. Научившись сотрудничать, они твердо полагаются друг на друга. Они моногамны и никогда не расторгают брачных союзов, хотя мужчин в группе больше, чем женщин. Как, возможно, было и у первых людей, тасадеи не имеют старейшины и не знают соперничества — каждый делает то, что у него получается лучше всего, а решения принимаются всей группой. Вот так живут тасадеи — в мире друг с другом и в гармонии с окружающей природой, — и наблюдавший их тихую жизнь антрополог считает, что они принадлежат к «самым кротким людям на земле».
Как видно, эти люди не пользуются практически ничем, что для современного человека является необходимым. У тасадеев для этого есть все возможности – они живут на Филиппинах, недалеко от экватора, в теплом круглый год климате. Они пользуются листьями пальм для одежды, но одежда служит тасадеям не для тепла, теплом их обеспечивает климат Филиппин. Видно, что женщина прикрывает импровизированной одеждой определенные части тела. Мы к этому вопросу еще вернемся, он не так прост, как кажется.
Если мы, обратив взор на север, туда, где холодно всегда, мы и там увидим людей. Люди на севере не могут жить без одежды. Эта одежда у северных народов есть вопрос жизни и смерти. Современные нам жители заполярья пользуются многими предметами, которые являются достижениями современной цивилизации. Тем не менее, эти жители вечного холода существовали там и сто и двести лет назад, беря в природе все необходимое и не нарушая природного равновесия. Их технические достижения были так же немалыми. На родной планете они жили как инопланетяне – жить можно только в искусственных жилищах, сохраняющих определенный температурный режим, выход за пределы теплого жилища – только в специальной одежде, обеспечивающей безопасный для жизни микроклимат. Но даже одежда позволяет находиться вне жилища только некоторое время. В музеях можно увидеть образцы такой одежды – не только теплой, но и легкой и непромокаемой. Отношение западного человека к таким людям всегда было и остается взглядом свысока. Западный человек всех неевропейцев всегда считал дикарями. Основанием для такого взгляда служили, в том числе, и многочисленные научные исследования, авторы которых пытались доказать, что неевропейцы – это не совсем люди. Собственно эти расистские взгляды европейцев и приходится оспаривать почти на каждом шагу.


Тем не менее, и белые европейцы, и негры и не приобщенные к цивилизации люди юга и севера – все мы принадлежим к одному биологическому виду. Все мы гости на этой планете, способные при скрещивании давать плодовитое потомство.
Этот вид образовался при определенных условиях около 10000 лет назад не на пустом месте. На территории Европы и СССР найдены многочисленные следы, говорящие о том, что длительное время современный человек жил одновременно с так называемыми кроманьонцами. Мало того, что он жил одновременно, они жили вместе, были биологически одним видом. Многочисленные находки переходных форм, которые трудно, а порой и невозможно однозначно определить как неандертальские или человеческие говорят о том, что человек современный и произошел от неандертальцев. Мало того, у современных людей можно встретить анатомические признаки, характерные для неандертальцев.
Современный человек имел предка, достаточно хорошо изученного археологами. Этот предок – кроманьонец – имел все признаки, характеризующие современного человека. Он умел говорить, жил в построенном им самим жилье, пользовался огнем, пользовался множеством предметов, ему было знакомо искусство. Рисунки кроманьонцев поражают наших современников. Таким образом, кроманьонец мало отличался от современного человека по образу жизни. Первые люди современного нам вида вели образ жизни кроманьонцев, и постепенно изменились. Современный человек имеет особенности анатомического строения, отличающие его от кроманьонца, при том, что образ жизни изменился не столько качественно, сколько количественно. Тип кроманьонцев не совсем исчез: кое-где он явственно проглядывает еще и в настоящее время, например в физическом облике населения некоторых местностей Франции (Неструх М.Ф. , 1970).
Отличие кроманьонцев от современного человека наиболее резко проявляется, прежде всего, в строении черепа. У современного человека жевательный аппарат развит слабее, чем у кроманьонца, есть отличия и в развитии надбровных дуг, и в строении мозгового черепа. Отличия же самого мозга за отсутствием сохранного материала многотысячелетней давности выявить труднее. Тем не менее, Кочеткова (1964) проведя исследования внутренней поверхности черепов, пришла к выводу, что некоторые размеры мозга и его частей имеют различия. Так, например, продольный размер мозга в среднем был у кроманьонца больше, чем у современного человека. Больше по длине у кроманьонца была и лобная доля (135 мм против 129 мм у современного человека). Таким образом, одним из противоречий антропогенеза и эволюционной теории Дарвина является относительное уменьшение размеров мозга у более позднего эволюционно вида.
Не менее интересно и происхождение кроманьонцев. Не вдаваясь пока в подробности обсуждения антропологами этого вопроса, отметим действительно интересный и таинственный факт – кроманьонцы появились около 40000 лет назад и практически в то же время исчезли неандертальцы.
Неандертальцы появились около 100 000 лет назад. Они пользовались огнем, жилищами, одеждой, но не говорили. Впрочем по этому поводу есть различные мнения. К этому вопросу мы тоже вернемся. Обратить же внимание следует вот на что. Когда обсуждаются темы антропогенеза, как то уходит из рассмотрения основной критерий. Я имею в виду определение человека. Что такое человек? Чем он отличается от животного? Какой критерий является наиболее существенным? Человек прямоходящий – это уже человек или еще животное? Использование орудий это чисто человеческая черта? Строительство домов и сложных гидротехнических сооружений доступно только людям? А как быть с бобрами? Что в современном человеке от животного, и какие анатомо-функциональные черты являются чисто человеческими, а какие мы можем встретить и у животного. Вера в бога доступна только человеку? Откуда это известно? Паломничество лососей к святым местам не уподобляет их людям? Опять же, откуда это известно? Кто-нибудь обсуждал этот вопрос с лососями? И что лососи ему ответили? Как они сами относятся к тому, что делают? Или человек не только расист по природе, но и убежденный враг всего живого? Пользователь, как теперь говорят, или еще грубее – потребитель. Общество потребления – это общество индивидов, убежденных, что главное дело на земле – перегон биомассы в удобрения.
Вопрос не праздный. Неандерталец жил относительно недавно, но австралопитеки появились на земле 5-6 млн. лет назад, а древнейшие орудия изготовленные австралопитеками найдены в Африке и датируются примерно 2 млн. лет. За такое время исчезли все некаменные артефакты, а они были? Или австралопитеки иного объекта приложения сил не нашли и творение начали сразу с камня. Можно ли называть австралопитеков людьми? Были ли они представителями, или предшественниками вида современного человека?
Получить ответы на все эти и многие другие вопросы, связанные с антропогенезом, значит получить базу для обсуждения будущего. На протяжении 20 века сделано достаточно много, для того, чтобы пересмотреть наши взгляды, дополнить исследования гениев 19 века, исправить ошибки, возникшие по недостатку знания и поставить на место негодяев, сознательно внедряющих ошибки в мировоззрение творцов будущего.
2.2 ЭВОЛЮЦИЯ ДВИЖЕНИЯ
Первые позвоночные появились на земле около полумиллиарда лет назад. Несомненным прогрессом перед лучевой симметрией предшественников позвоночных было два момента – во-первых выделение головного конца, на котором впоследствии сконцентрировались органы чувств, и центральный скелет, придающий прочность телу, но не защищающий его от внешних воздействий. Не имея защиты на поверхности тела, позвоночные вынуждены были придавать большое значение сигналам внешней среды, получаемым органами чувств. Для выделения значимых для жизни сигналов, распознавания опасно-неопасно, привлекательно-непривлекательно, была нужна постоянно развивающаяся нервная система, в данном случае трубчатая. Надо отметить, что на момент появления первых позвоночных животная жизнь на земле существовала давно, а дышащие кислородом животные существовали к тому времени уже полмиллиарда лет. Эти животные ангельской кротостью не отличались, а первые позвоночные не были крупными, и в хищничестве не были лидерами.
Дальнейшая эволюция позвоночных происходила по линии древние рыбы – амфибии – пресмыкающиеся – млекопитающие.
В этом ряду происходило совершенствование всех систем. Совершенствовался опорно-двигательный аппарат, прежде всего, скелет. От несегментированного скелета ланцетника к четко выраженным сегментам скелета костистых рыб, затем появление и развитие скелета конечностей. Мышцы опорно-двигательного аппарата так же претерпели изменения, но менее заметные.
Очень сильно изменилось снабжение организма кислородом – от жабр и кожного дыхания у рыб и амфибий, до легких у более поздних видов. Кровь все более эффективно переносила газы, осуществляя их обмен и, тем самым энергообеспечение организма, а сердечно-сосудистая система стала замкнутой, с четким выделением двух кругов кровообращения.
Но наибольшее изменение претерпела нервная система. Основное направление эволюции позвоночных приобрело в связи с этим название «цефализация». Под этим термином подразумеваются две стороны явлений:
1). прогрессирующая утрата самостоятельности и функциональное обеднение задних отрезков центральной нервной системы – спинного мозга и
2). постепенное перемещение вперед наиболее прогрессивных центров мозга. То есть непрерывное усиление роли головного мозга на каждом этапе эволюции. Каждое новое эволюционное приобретение в ряду позвоночных сопровождалось как объемным (или весовым) приращением головного мозга относительно соответствующих параметров тела, так и его качественным преобразованием. Это преобразование проявлялось во все более совершенном управлении телом. Кроме того, центральная нервная система забирала все больше кислорода, потребляемого организмом.
Цефализация проявила себя в том, что задние отделы ЦНС имели все меньшее значение, по сравнению с передними. Если у млекопитающих спинной мозг располагается по всей длине туловища, то у взрослого человека спинной мозг едва достигает 2-го поясничного позвонка. Головной же мозг, который делится на отделы – задний мозг, средний мозг и передний мозг – развивается в процессе эволюции так же неравномерно. В ряду от рыб до человека в наибольшей степени развивается передний мозг. Он образует два полушария, покрытые серым веществом, но и в этом отделе происходит все та же цефализация. По сравнению с млекопитающими у человека не только сильнее развита кора полушарий головного мозга, но и среди отделов коры наиболее развиты лобные доли.
На рис 3 и 4 из учебника биологии приведены схемы головного мозга у различных животных в ряду позвоночных. К конечному мозгу имеет отношение то, что на всех рисунках обозначено цифрами 1 и 8. Хорошо видно, что в наибольшей степени полушария головного мозга в наибольшей степени развиты у млекопитающих. Это и есть иллюстрация цефализации в достаточном для нашего рассмотрения виде. рис 3,4
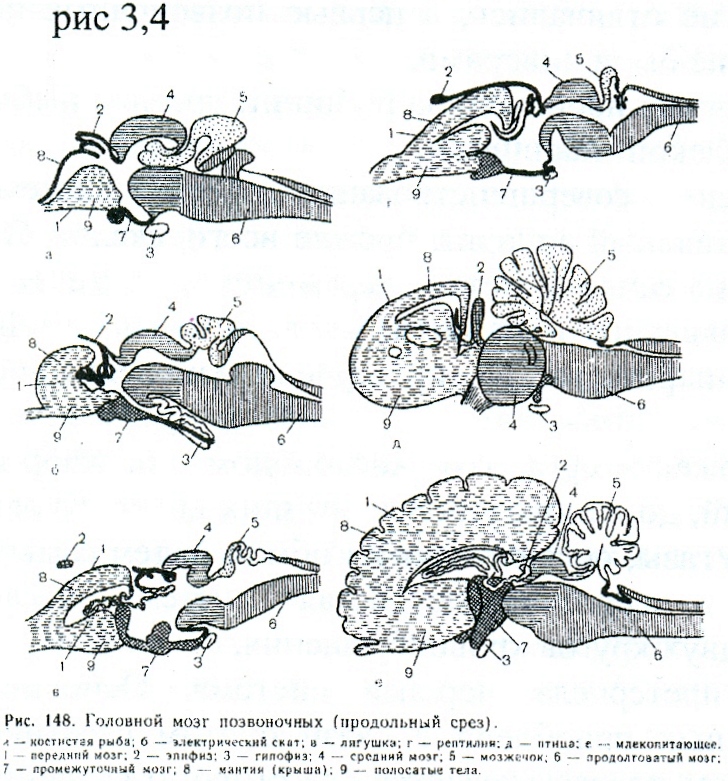
Если взять за отправную точку изображение головного мозга млекопитающего на нашем рисунке, то сравнение человека и млекопитающих приведено на приведенных рисунках (из Неструх М.Ф.1970).
 рис. 5
рис. 5
На этом рисунке представлены фотографии головного мозга человека и млекопитающих, вид сверху, в реальном соотношении размеров. Не очень правильный для понимания ракурс, т.к. не видно нижележащих отделов головного мозга. Кроме того, необходимо учесть размеры тела животных (рис. 5 — слева). Видно, что головной мозг у слона почти вдвое больше, чем у человека. Но если принять во внимание, что вес человека редко достигает центнера, а вес слона достигает нескольких тонн, то становится понятно, что на единицу веса тела у слона приходится в несколько десятков раз меньше нервной ткани, чем у человека.
Не менее интересно сравнение головного мозга человека и приматов. Рисунок справа.
Каким путем шло развитие человека? Почему мы сравниваем его головной мозг с головным мозгом приматов. Дело в том, что как принято говорить, человек произошел от обезьяны. Это упрощенное выражение, приписываемое Дарвину. На самом деле Дарвин говорил об общих предках обезьяны и человека.
В 20 веке было сделано огромное количество находок, на основании которых были сделаны следующие выводы.
В Африке и Азии на протяжении от 2 млн. лет назад и примерно до 100 000 лет назад жило существо, названное учеными «австралопитек». Разновидности австралопитека названы человек прямоходящий и человек умелый. Эти две разновидности мало различимы по костным окаменелостям, сохранившимся до наших дней. Главное различие в том, что человек прямоходящий не делал каменных орудий, а человек умелый делал их в большом количестве. Впрочем, речь идет только о каменных изделиях. Совершенно не осталось никаких следов изделий из других материалов. Кроме того, известно, что примерно 800 000 лет назад австралопитек пользовался огнем. Пользование огнем австралопитеком было ограничено – очень мало находок. Известно, так же, что австралопитек строил жилища, используя для этого большие камни и деревянные детали.
На самом деле об австралопитеке известно немало. Для интерпретации таких данных нам необходимо сделать небольшой экскурс работы совершенно иного направления.
2.3 ФИЗИОЛОГИЯ ДВИЖЕНИЯ
В числе таких трудов, в первую очередь следует назвать работы Н.А. Бернштейна в области физиологии движений. Не оцененным антропологами по достоинству его труд оказался по причине его огромного значения в достаточно специфической области – физиологии трудовой и спортивной деятельности. В связи с этим, в настоящей работе нам придется обильно цитировать работу Н.А. Бернштейна «Физиология движения и активность», тем более, что автор сам придавал большое значение антропогенетическим аспектам в этой работе.
«Если бы мы захотели представить прогресс (физиологических функций) графически, то для функции дыхания или обмена он изобразился бы линией, вряд ли существенно отклоняющейся — в пределах филогенеза теплокровных — от параллельности с осью абсцисс. Кривая развития психических функций имела бы все основания выглядеть на подобном графике очень круто восходящею кверху; но, к сожалению, мы имели бы объективное право нанести на чертеж только ее самую правую (самую верхнюю) площадку, относящуюся к человеку. Вся остальная кривая осталась бы в области гипотез из-за полного отсутствия объективного материала, касающегося животных, несмотря на всю героику объединенных усилий зоопсихологов, бихевиористов и кондиционалистов. И только для двигательных отправлений мы можем вполне реально и объективно построить полностью их эволюционную кривую, круто восходящую к правому концу графика и далеко обгоняющую темпами своего развития сам по себе весьма не медленно эволюционирующий их морфологический центрально-нервный субстрат. Уже одно это обстоятельство делает физиологию движений интересной для психолога и невролога» — пишет Н.А., а мы добавим – и для антрополога. Н.А. Бернштейн выделяет уровни регуляции движений. В соответствии с его трудами каждый новый эволюционный уровень получает новый аппарат для управления движениями.
Каждый уровень содержит определенный набор движений человека. В совокупности уровни управления движений образуют рекапитуляцию всей истории животных движений, начиная от перистальтикоподобных движений кольчатого червя.
Такая рекапитуляция имеет преимущества перед нейроморфологией, поскольку воспроизводит филогенез не в статике и не в символике гистологических обликов нервных ядер, ничего не говорящих о своей функциональной сущности, а в динамике, в движениях, доступных точным сравнениям по содержанию, смыслу, оформлению у современных нам представителей всех ступеней филогенетической лестницы. У самого дна, глубоко в толще двигательных координаций человека, мы находим древнейшие палеокинетические координации, отошедшие у высших позвоночных в удел вегетативным отправлениям: перистальтические движения кишечника, стрикционно-дилятационные движения в сосудистой системе и т.п. Поднимаясь выше, мы встречаемся с первичными, самыми элементарными и по структуре и по определяющей их афферентации неокинетическими координациями».
Здесь необходимо остановиться подробнее. Дело в том, что при изучении нервной системы, студенты опираются на учение Павлова о рефлекторной организации функций нервной системы. Несомненная доказательность рефлекторной теории, и авторитет Павлова сыграли злую шутку в первую очередь над изучением физиологии ЦНС студентами, а следовательно и в последующем врачами и учеными. Дело в том, что сколько бы мы ни изучали рефлексы, полное их строение останется для нас тайной, пока мы не станем использовать иную более плодотворную концепцию. Такой концепцией и является концепция Н.А. Бернштейна. В основе ее лежит понятие «афферентный синтез». Афферентный синтез создает картину мира, доступную для данного отдела нервной системы. В рамках этой картины мира, или афферентного синтеза и происходит управление двигательной активностью организма.
В ряду позвоночных – от водных форм жизни, через пресмыкающихся к теплокровным и млекопитающим происходит ступенчатое усложнение систем управления движениями. Этих уровней Н.А. Бернштейн насчитывает 5. Он обозначает их буквами – А (тонусы), В (синергии), С (локомоции), Д (предметные действия), Е (речь). Начиная с кортикальных уровней, развитие каждого последующего уровня возможно только с кардинального усложнения картины мира, или афферентного синтеза.
Полное развитие любого уровня возможно только при появлении хотя бы слабого, но более высокого уровня регулирования, а соответственно и более сложного афферентного синтеза. Если у организма нет более высокого уровня, если данный уровень регулирования движений является наивысшим, то он никогда не разовьется в полной мере, не раскроет всех своих возможностей.
Реально каждый высший уровень работает только при освоении новых двигательных навыков. По мере освоения движений происходит их автоматизация, т.е. освоенные движения управляются на более низком уровне, чем это происходило при освоении двигательного навыка. На самом деле при освоении двигательных навыков происходит поиск сенсорных коррекций для афферентного синтеза на более низком уровне, или, иначе говоря, вышележащие отделы мозга обучают нижележащие отделы, после чего считают свою функцию выполненной, и в работу, по возможности, не включаются. Чем выше отдел головного мозга, тем больше он стремится не работать, тем труднее он включается в работу. Соответственно, по мере освоения двигательных навыков, управление ими переходит на более низкие отделы головного мозга.
В формировании двигательных навыков четко выделяют несколько фаз:
1. Стадия генерализации (иррадиации возбуждения). На этой стадии под генерализацией понимается максимальное распространение возбуждения в коре головного мозга. При этом функционально расширяются ассоциативные связи нейронов разных зон. Это облегчает поиск решения двигательной задачи. Надо понимать, что кора головного мозга ищет приемлемое решение данного вида двигательных задач, и даже при автоматизации действия каждое повторение есть поиск и использование алгоритма решения. Задача может быть одна и та же, но решение каждый раз с учетом конкретных условий новое. На этой стадии нет согласованности двигательных и вегетативных реакций. Массированный поток афферентной импульсации от мышц, внутренних органов затрудняет выделение информации от основных рабочих мышц. В сочетании с генерализацией возбуждения в коре головного мозга происходит расширение афферентного поля и возможностей для нового афферентного синтеза. Расширенное афферентное поле позволяет поиск новых сенсорных коррекций, а в них то и нуждается новый двигательный навык.
2. Стадия концентрации возбуждения в необходимых для действия центрах и подавление – в лишних. Энергозатраты на этой стадии снижаются, но автоматизации еще нет, то есть управление происходит с участием коры головного мозга. Обучение подкорковых центров не закончено, и при даже небольших изменениях условий – подкорковый центр начинает требовать от коры уточнения инструкций, как следствие возникает иррадиация возбуждения в коре и потеря координации.
3. Стадия стабильности и надежности. Подкорковые центры уже обучены действиям, сознательный контроль коры прогрессивно снижается, т.е. произошла автоматизация навыка. Афферентный синтез происходит на основе строго отобранной информации, и новые посторонние внешние раздражители не вносят помех. Более того, на этом этапе они подкрепляют доминанту, управление передается все ниже.
Низшим субкортикальным уровнем является уровень А. Функции уровня А – это функции спинного мозга, перенесенные в головной мозг. На этом уровне держится управление тонусом мышц. Движения этого уровня – дрожь от холода, стучание зубами, вздрагивание, а в области произвольной моторики – быстрые ритмические вибрационные движения (фортепианные вибрато, вибрации левой руки скрипачей, обмахивания веером), движения, связанные с принятием и удержанием определенной позы (полетная фаза прыжка в воду). Статически – хваточные позы.
Синергии и штампы (Б) – таламо-паллидарный уровень.
Уровень Б обогащает моторику позвоночных по сравнению с уровнем А. Он так же, как и А, является субкортикальным и соответствует анатомически таламо-паллидарной системе.
Зрительный бугор — таламус производит афферентный синтез, а бледный шар – управление движениями уровня. Этот уровень имеет место уже у рыб, у них он наиболее развит, имеет важное значение у земноводных. Для рыб характерны плавные движения, охватывающие все тело, представляющие собой что-то вроде перистальтических волн. Эти движения – плавников, хвостовых перьев и т.п. – не прекращаются ни на минуту даже и при спокойном стоянии рыбы на одном месте. В инвентаре движений почти все 100% составляют плавательные локомоции, оформляемые как крайне обобщенные, охватывающие всё тело синергии. Движения по большей части неторопливы, округлы и ритмичны.
У позвоночных сухопутного образа жизни этот уровень приобретает особое значение. Позвоночные на земной поверхности перемещаются при помощи конечностей. Конечности представляют собой многозвенные рычаги. Каждый из этих рычагов соединен с соседним при помощи суставов. Подвижность в суставах ограничена их формой. На каждый сустав действуют мышцы как минимум в двух направлениях – если одна группа мышц осуществляет сгибание в суставе, то другая группа мышц будет осуществлять разгибание в этом суставе. Движение в каждой мышце (сокращение и удлинение) усложняется тем, что это движение не подобно движению поршня в цилиндре. Для более близкого подобия концы поршня и цилиндра надо прикрепить к рычагам не непосредственно, а через пружины (аналоги сухожилий). Всю сложность управления конечностями для Вашего мозга, вы можете себе представить таким образом. Привязав к кольцу лыжной палки два резиновых жгута, уприте рукоять палки себе в живот, и, взяв в руки концы жгутов, попробуйте написать острым концом лыжной палки любую заранее заданную фигуру. Вся сложность задачи предстанет перед вами, и прежде всего, обуздание реактивных сил, возникающих при каждом движении. Необходимость локомоций при помощи конечностей привела к тому, что основной уровень управления движениями у рыб, обеспечивающий у них перемещение в пространстве, взял на себя роль мощного интегрирующего, и, главное, синергирующего аппарата ЦНС. Синергии подразумевают согласованное управление напряжением мышц-антагонистов.
Афферентный синтез этого уровня осуществляется в таламусе. Сюда поступает очень специфическая информация. Здесь не учитываются данные от дистантных рецепторов – зрительных и слуховых. Сюда поступает информация только от рецепторов, связанных с телом. Это рецепторы кожной чувствительности, рецепторы мышц, суставов, связок, костей, и рецепторы состояния внутренней среды организма. Здесь формируются целостные телесные ощущения (голод, холодно-тепло, удобно-неудобно, жажда и т.п.). на основании такого афферентного синтеза происходят сенсорные коррекции уровня Б.
Управляющие движениями импульсы исходят от базальных ядер называемых бледный шар. Это движения всего тела лишь с ним одним связанные, безотносительные к чему – либо находящемуся вовне, движения как самоцели, корригируемые лишь по линиям своей внутренней гармонии и организованности.
Три важнейших качества этой системы:
1). обширные мышечные синергии;
2). способность столь же стройно и налажено вести движение во времени, обеспечивать правильные чередования, например, перекрестные чередования движений конечностей при локомоциях, объединять в общем ритме;
3). наклонность к штампам, к чеканной повторяемости движений.
Самостоятельно этот уровень ведет движения выразительной мимики, пантомимы и пластики, т.е. совокупность не символических, а непосредственно эмоциональных движений лица, конечностей и тела. Уровень синергий не приспособлен к использованию ни зрительного ни слухового контроля и управления.
Почти не выходя за пределы уровня протекают многие движения вольной бесснарядовой гимнастики: наклоны корпуса, изгибы, откидывания тела, пластико-ритмические движения.
Сюда же относится группа полунепроизвольных движений – потягивания всем телом, расслабления членов, движения ласки (объятия, поцелуи), привычные монотонно-машинные движения. Все движения этого рода плавны, гармоничны, грациозны даже у неграциозных людей. Именно этот уровень больше других приспособлен к обузданию реактивных сил и к созданию экономичных траекторий динамически устойчивого типа. Слабость уровня синергий не в качестве или количестве афферентаций а в их односторонности. Ему не хватает дистантных рецепторов и их синтеза, ведущего к овладению внешним пространством. Его система координат привязана не к окружающему эвклидову пространству а к собственному телу. Этот уровень слишком интравертирован. Он не годится ни в штурманы, ни в пилоты движения, он – бортмеханик.
Фоновая роль уровня синергий огромна и не уступает роли уровня А. будучи не в состоянии из-за отрыва от телерецепторов и слабости связей с вестибулярной системой и мозжечком вести много самостоятельных движений, уровень Б берет на себя черновую технику сложного движения, если другой вышележащий уровень обеспечивает его приспособление к внешнему миру и предметам.
Существенна роль синергий в автоматизации двигательных навыков. Автоматизация сводится к передаче ряда технических координационных компонент двигательного акта в нижележащие, не сознаваемые в данный момент уровни (осознается всегда только ведущий уровень совершаемого движения). При этом в уровне Б вырабатываются новые двигательные комбинации, или вырабатывается умение использовать имеющиеся готовые автоматизмы из фондов этого уровня. Иначе говоря, более высокие уровни учат уровень Б как вести себя в том или ином случае. Будучи обученным, уровень работает без участия сознания.
Усложнение двигательных задач и обогащение координационных ресурсов совершенствуются по двум линиям: за счет эволюции рецепторики и за счет эволюции центральных замыкательных систем.
Во-первых, двигательные задачи усложняются. Усложняется мир, вместе с ним возрастает разнообразие реакций, требующихся от организма. насколько аэродинамический полет птицы сложнее почти полностью гидростатического плавания рыбы. Молодая отрасль проворных теплокровных млекопитающих победила тугоподвижных юрских завров именно своею более совершенной моторикой. Это произошло за счет эволюции рецепторики.
Во-вторых, все возрастает количество разовых задач, непредвиденных в генетически заложенных программах. ЦНС приспосабливается к сложнейшим условиям, перестраивается практически мгновенно, возрастают способности к накоплению индивидуального опыта. Это – за счет эволюции центральных замыкательных систем.
Эволюция рецепторики — во первых переслоение тактильной чувствительности – дистантной. Появление новой формы проприоцепторики — геометрической, воспринимающей позы и скорости и возглавляемой неолабиринтом полукружных каналов. На фоне древней – проприоцепторики тропизмов, возглавляемой отолитовым аппаратом (палеолабиринтом) и приспособленной к оценке давленией, напряжений и усилий, к ориентировке в поле тяготения.
В третьих – все более развивается система телерецепторов :
1). переход от однонейронной таламической схемы к схеме кортикальной афферентации;
2). переход от узловой системы нервноклеточных ядер к двухмерной экранной схеме, характерной для коры полушарий;
3) усложнение синтетических сенсорных полей. Эти сенсорные синтезы, в которых сырые рецепции отдельных органов чувств сливаются вместе с памятью индивидуального опыта и направленные на координацию движений и действий, стимулируют развитие центральных замыкательных систем не менее, чем телерецепторы. Филогенетическое формирование этого ряда усложняющихся полей сопряжено с ростом удельного веса памяти.
Рост направляющего влияния телерецепторики на эволюцию двигательных функций объясняется еще и тем, что ею были вызваны сложные двигательные формы (локомоции) а эти последние потребовали применения сенсорных коррекций. Таким образом, движения стали опираться на рецепторику, тогда как у древнейших форм, наоборот, рецепции вытекали из движения.
Возрастание удельного веса разовых реакций, c опорой на индивидуальный опыт, связано с эволюцией коры полушарий. Развитие последней переводит ЦНС из подчиненного положения в положение возглавляющей и направляющей.
Ход филогенетического развития строения центральной нервной системы состоит в качественном обрастании ее новыми образованиями, не имеющими гомологов в предшествующих этапах филогенеза, и представляющими собой надстройки не один или больше нейрональный этаж на пути следования нервного процесса. Этот принцип приводит к неминуемой скачкообразности развития ЦНС уже из-за дискретности нейронной схемы: усложнение рефлекторной дуги возможно не иначе, как на целое число новых вставочных нейронов.
Возникновение в филогенезе очередной новой мозговой надстройки знаменует собой биологический отклик на новое качество или класс двигательных задач. При этом обязательно появляется новое синтетическое сенсорное поле, а тем самым и появление возможности нового класса движений, качественно иначе построенных и иначе управляемых, чем те, что были раньше. Вся совокупность морфологических и функциональных сторон, характерных для такого нового класса движений обозначается как очередной уровень построения движений и двигательных координаций.
Более новые в филогенезе уровни становятся все более тесно связаны с телерецепторикой и надстроенными над ней обобщающими системами в коре головного мозга, все более экстемпоральными, т.е., пригодными для осуществления разовых решений; все более синтетичными, т.е., опирающимися на сложные психологически организованные сенсорные поля, все более богатыми памятью, т.е., обогащаются индивидуальным опытом.
Пирамидно-стриальный уровень С.
Афферентный синтез формирует образ пространственного поля, движения в этом поле характеризуются как локомоции (перемещения). Н.А. Бернштейн так характеризует этот уровень:
«Первичные сенсорные станции в коре — зрительные поля, постцентральная извилина, слуховые и вестибулярные поля, новый мозжечок.
Элементы организации пространства имеются и на уровне А, где они проявляются в виде тропизмов тяготения, выступая как почти беспримесная полярность верха и низа. В таком именно виде мы встречаемся с пространством, как координационным фактором, и у насквозь палеокинетических бесскелетных беспозвоночных, где поля тяготения определяет и стойко выдерживаемое расположение тела в пространстве, и распределение мышечного тонуса. В первом неокинетическом, централизующем уровне В пространство проявляет себя в виде системы координат собственного тела, обобщенной в единой системе местных знаков, но еще не спроецированной на внешнее дистантное окружение.»
Пространственное поле уровня С не есть ни ощущение, ни их сумма. Пока оно формируется, в нем участвуют и зрительные, и глазодвигательные ощущения, связанные с аккомодацией и стереоскопическом зрением, и осязательные ощущения и проприоцепторика тела, и вестибулярные ощущения тяготения и ускорений.
Пространственное поле полностью соотнесено с внешним миром и освобождено от неотрывной связи с собственным телом. Оно обширно, несдвигаемо, гомогенно. Обширность во многом обеспечивается телерецепторами. Слепые его формируют локомоцией с осязанием. Для объективации пространственного поля важно ощущение его несдвигаемости. Когда мы передвигаемся, мы не только знаем, но и ощущаем что перемещаемся мы, а пространство с наполняющими его предметами неподвижно, хотя все рецепторы говорят нам об обратном.
Пространственное поле гомогенно и апериодично, т.е. однородно во всех частях и не содержит в себе никаких элементов чередования или циклической повторяемости, которых так много в координатной системе собственного тела.
Оно метрично и геометрично, оно включает в себя точную оценку протяжений, размеров, форм, входящих в качестве существенных признаков и в движения, выполняемые на этом уровне: это область точности и меткости. Геометричность поля характеризует в наибольшей мере верхний, кортикальный подуровень С2, проявляясь в соблюдении геометрической формы и геометрического подобия. Это самое объективное из пространств, дальше кверху, в уровне действий, оно эволюционирует в сторону схематизации и, выигрывая в смысловой упорядоченности, теряет в объективности.
Пространство С не пустое: оно заполнено объектами, имеющими размер, форму и массу и силами, действующими между этими объектами. Эти силы исходят от внешних тел и привязаны к ним, а не к нашему телу.
Этот уровень – потолок моторных возможностей высших рептилий, птиц и у низших млекопитающих и у человека до второго года жизни, продолжает преобладать у него в течение всего детства.
Движения уровня пространственного поля имеют ясно выраженный целевой характер: они ведут откуда-то куда-то и зачем-то. Движения имеют начало и конец, приступ и достижение, замах и бросок или удар. Движения в пространственном поле всегда переместительны, если их внешнее оформление иногда циклично в силу устройства наших конечностей, то по смысловой структуре они апериодичны, как и само пространство. Переместительное движение по своей сути предполагает прилаживание к условиям того внешнего пространства, в котором оно протекает.
Управление движениями происходит из двух пунктов: полосатое тело (подуровень С1), и моторная кора прецентральной извилины (подуровень С2).
Особенностями управления на уровне С1 — приспособительность по ходу процесса, точность в деталях движения. Обведение контура фигуры. Непроизвольные коррекции движения в соответствии с локальным характером пространственного поля.
Эта ступень начинает развиваться у сухопутных амфибий, четко представлена у рептилий и достигает кульминационной точки своего развития у птиц. Преобладание полосатого тела обеспечивает этим животным передвижение уже не туловищного, а конечностного стиля. Очень высокой ступени развития достигает владение равновесием. Статика уравновешенной рыбы не составляет проблемы для нее; здесь напротив, вырабатывается целая система статокинетических регулирующих механизмов. Сохраняя все фоновые синергетические возможности, обеспечиваемые уровнем Б, эти животные располагают уже средствами для их затормаживания и регуляции. Они способны застывать неподвижно и делать порывистые броски. Движения туловища и головы имеют своеобразный характер, вязкий, тестообразный, сменяясь временами на восковую, статуеобразную неподвижность, совершенно чуждую млекопитающим.
Наряду с усложнением и увеличением разнообразия локомоции имеет место и возникновение целого ряда движений других категорий, иногда создающих иллюзию уровня Д (действий) (например, витье гнезд). Птице доступны движения, не встречаемые у рыб: движения туалета, смазывание перьев жиром, кормление птенцов и т.п. Здесь впервые появляются выразительные движения и звуки: мертвенные крики лягушек сменяются песней, возникает и танец. При этом пластичность и способность к построению новых двигательных комбинаций очень низки.
На уровне С2 – выраженный целевой или финальный характер, движение проецируется на его конечную точку, с установкой на точность. Этот подуровень индифферентен к траекториям, полностью концентрируется на конечном пункте.
Здесь – господство уровня пространственного поля С во всем его объеме, сопряженное с возникновением и развитием пирамидной эффекторной системы и наступающее впервые у млекопитающих. Не теряя ничего из координаций, приобретенных на нижележащих уровнях животные приобретают большое количество однократных, не стереотипных целевых движений нападения, охоты, скачка и т.д. Все более возрастает пластичность, обучаемость, способность к импровизации новых двигательных комбинаций. Резко разнообразятся движения туалета и самообслуживания. Обильными становятся игры в компании, педагогические действия и показы, отсутствующие у более древних классов позвоночных. Наряду с выразительными звуками и интонациями появляется мимика и выразительные движения. Движения становятся эластичными, пружинными, покой перестает быть абсолютным, насыщаясь то настораживательными движениями, то привычными ритмическими покачиваниями хвоста, то иной непроизвольной двигательной мелочью. Постепенно начинают появляться истинные смысловые цепи, зачатки предметного уровня.
Виды движений в пространственном поле:
Локомоции: ходьба, ползание, плавание, коньки, лыжи, велосипед, гребля, прыжки с разбега и с места, ходьба с тягой и толканием.
Нелокомоторные движения: акробатика, гимнастика. Перемещение вещей в пространстве: взятие, схватывания, ловля летящего предмета, передвигание, перенос, доставание, наматывание, преодоление внешних сил – подъем тяжестей, натягивание лука и т.п.
Баллистические движения: на силу (толкание, метания, удар молотом, рывок штанги) на меткость (метания, жонглирование, удар по зубилу, укол штыком, все эти движения требуют зрительного контроля).
Прицеливания и установочно-выжидательные движения.
Подражательные и копирующие движения: имитация, срисовывание, изображение жестами, пародирование.
Овладение внешним пространством требует справляться с внешними силами, сюда относятся силовые движения: поднимание тяжестей, баллистические движения.
Срисовывание фигуры. Коррекции движения произвольные.
Теменно — премоторный уровень действий (Д) (предметные действия).
Все координации и значительная часть движений, описанные в предыдущих главах принадлежат и животным. Более того, есть виды, обладающие долее высоким развитием пространственного поля, чем у человека. Животные лучше бегают, плавают, зорче, метче и точнее нападают. Кульминационные точки развития по этому уровню уже пройдены, правда, списки движений уровня пространственного поля у человека богаче. Но этот уровень еще не совсем кортикальный.
Уровень Д почти монопольно принадлежит человеку, именно в нем строятся главнейшие фоны речевых и графических координаций, и этот уровень явно далек от своей кульминации. Он едва представлен у лошадей, собак, слонов. Даже у высших обезьян процент его очень мал, и к нему прибегают только после неудачи решить задачу на более привычных нижележащих уровнях. При апрактическом нарушении страдает не координация двигательного акта, а его реализация. Больному покорны все органы, но он сам не умеет сделать посредством их ничего, что выходило бы за пределы элементарных двигательных комбинаций, присущих и животным. Теряется способность к приобретению или восстановлению умений и навыков, превышающих эти элементарности.
Необходимая зона уровня действий – левая теменная область.
Ведущая афферентация есть предмет. Психологически образ предмета представляет собой результат гораздо более глубоких обобщений и гораздо более сложной синтетической связи между сенсорными и мнестическими составляющими, нежели синтез, называемый пространственным полем.
Ведущий мотив на уровне действий не предмет сам по себе, а смысловая сторона действия с предметом – как объектом или орудием.
Здесь слиты воедино 3 понятия – предмет (верхняя теменная долька) смысловая структура действия (нижняя теменная долька) и его двигательный состав (премоторная кора).
Предмет может восприниматься как объект на который направлено действие, как орудие воздействия, как символ (шахматная фигура, буква).
Большой интерес для учения о координации представляет особенность предметных действий, выражающаяся в ведущей роли, захватываемой предметом, и свидетельствующая о глубоких ускользающих от сознания перешифровках. Когда предмет фигурирует в качестве инструмента при высокоавтоматизированной привычной работе, он прямо переживается исполнителем действия как органическая часть собственного тела, вплоть до иллюзии активного ведущего управления движениями, как бы исходящего из этого инструмента.
На уровне Д вместо геометрической формы появляется схема, т.е. метрические, размерные соотношения заменяются топологическими, качественными соотношениями. Пространство предметного уровня теряет в конкретности, но выигрывает в упорядочении, выделении существенного по сравнению с уровнем пространственного поля. В нем выделяются и организуются качественные понятия замкнутой и разомкнутой фигуры, представления «над» «под» «вне» «внутри» «между» и т.п., происходит категориальная организация пространства, т.е. возобладание топологической смысловой схемы над геометрической формой.
В геометрическом образе мы можем различать его топологию и его метрику. Топологическая схема не имеет размера. Топологически все ботинки одинаковы, но топологический ботинок может не подходить Вам метрически.
Автоматизация движений состоит в переключении ряда координационных фоновых компонентов движения в нижележащие уровни, что связано с их переключением на другие афферентации. При автоматизации на уровне синергий зрительный контроль заменяется на осязательно-проприоцептивный. При передаче с уровня действий на уровень пространственного поля автоматизация сопровождается переходом с ментальных схем на зрительные коррекции (неумеющие рисовать рисуют не натуру а идеограммы).
Пространство Д строится по принципу топологической схемы, а не геометрической формы. Все вносимое этим уровнем в движения топологично, а не метрично. Не метричны: снимание шляпы, закуривание папиросы, изображение домика, человека, завязывание узлов, набирание кружкой воды. Вопрос об успешности движения решается по качественному результату. Предметы на этом уровне геометричны — топологичны и метричны.
Для каждого уровня характерно иное восприятие времени. Время на уровне синергий – ритм, на уровне пространственного поля – момент, синхронность длительность, скорость. На уровне действий – причинная последовательность (до и после).
Пространство на всех уровнях связано с афферентацией. Из афферентации вырастает пространство, из пространства – предмет, из предмета — объектные понятия. Таким образом, эволюционная цепочка афферентация – пространство – вещь – объект от начала до конца связаны с задними отделами мозга.
Время в ЦНС связано с эффекторикой. На уровне синергий оно влито в состав движения, воплощая его ритмовую динамику. На уровне пространственного поля – скорость, темп. На уровне предметного действия время претворяется в смысловую связь и последовательность активных действий по отношению к объекту.
Из эффекторики вырастает субъективное время, из времени – смысловое действование, из него – поведение, из поведения – субъект.
Эволюция движение – время – субъект – связана с развитием лобных долей.
Уровень синергий для предмета – фон. Явно предмет появляется только на уровне пространственного поля:
1). как пункт приложения сил, материальная точка;
2). как смысловые отношения;
3). как действия.
Движения в уровне предметного действия — смысловые акты, не столько движения, сколько элементарные поступки, определяемые смыслом поставленной задачи. Одеться, забить гол, очинить карандаш.
Упражняемость действий, автоматизируемость, появление высших автоматизмов – предметных навыков – сноровок, при том, что сноровки не являются предметными действиями. Характерное свойство автоматизмов — приобретенность.
Повреждения в нижне-теменных отделах полушарий – нарушают смысловую структуру сложных двигательных актов или действий, а очаги в премоторных полях – нарушения в двигательном составе.
Все движения предметного уровня – смысловые акты – непосредственно не витальны. Это не еда, не тропизм за едой и самкой, не обеспечение функции организма. Это всегда промежуточная цель, отвлечение от окончательной цели. Поскольку окончательная цель всегда витальна, она определяется древней, лимбической корой. Лимбическая кора делегирует свои функции лобной коре для совершения всех промежуточных действий. Тем не менее, контроль лимбической коры не прекращается, более того, снятие такого контроля приводит к совершению действий не нужных.
Уровень Е мы рассмотрим немного позже.
Таким образом, мы видим, что основная разница между человеческим поведением и животным проходит в степени развития уровня Д – предметных действий. Этот уровень наиболее полно представлен у человека, у животных он появляется у млекопитающих и у них, даже высших он представлен фрагментарно.
Уровень предметных действий позволяет практически осознанно пользоваться законом причинности – время на этом уровне воспринимается как до и после (не момент, не длительность, не повторяемость и не ритм). Закон причинности – это уже понимание смысла происходящего.
В то же время, уровень предметной деятельности даже наиболее близким к людям, нашим ближайшим родственникам доступен не всегда. Для действия на уровне предметного человекообразные обезьяны должны убедиться в невозможности решить задачу другими способами. Разрешение этой ситуации должно быть жизненно необходимо, иначе уровень остаётся недоступным. Если же ситуация способна привести кору головного мозга животного в состояние предельной активности, то необходимый синтез становится возможным, животное решает задачу на уровне Д. Ряд повторных решений приводит к автоматизации действия, и в последующем, при повторении ситуации животное действует более низкими уровнями, т.е. просто перемещает себя или предметы не приходя в состояние чрезмерной активности, какое единственно могло включить афферентный синтез предмета. Нет нужды у братьев наших меньших постоянно находиться на нашем уровне понимания ситуации. И никогда не было. У наших же гипотетических предков подобные ситуации были у многих поколений непрерывно в течение всей жизни. Иначе на уровень Д эволюционно не подняться. Как это могло выглядеть.
Моделируя ситуацию необходимо помнить, что уровень предметных действий, для закрепления в генетическом аппарате должен был быть востребован долго, постоянно, и задолго до формирования уровня Е (речи). То есть наш гипотетический предок австралопитек, практически все изменения в строении тела и в поведенческих реакциях произвел в состоянии постоянно действующего фактора, который и стимулировал работу его высшего уровня управления телом. Все наши предположения, помимо прочего, должны включать в себя и необходимость для нашего предка думать на пределе его возможностей.
И для появления первых обезьян эта сторона вопроса совершенно не представляет трудностей. Лазание по деревьям представляет собой довольно сложный вид локомоций. Перемещение в трехмерном пространстве предъявляет высокие требования к уровню локомоций. Само по себе перемещение не на плоскости, а в объеме требовало учета расположения опорных точек для конечностей на всем протяжении маршрута. Если конечный пункт маршрута намечался на уровне С2, то коррекция каждого шага происходила на уровне С1. каждый шаг был отличен от другого, оценка расстояний от одной опорной точки до другой, опорные характеристики этих точек, возможности конечностей для их использования – все это вносило серьёзнейшие изменения и в строение коры и базальных ядер головного мозга, и в мозжечок, и в работу зрительного анализатора. Каждая точка опоры рассматривалась в точном соотнесении с параметрами конечностей, возможностей надежного захвата, с учетом прочности именно этой ветки, ее способностей выдержать вес тела.
Трехмерное пространство локомоций и рассмотрение в трех измерениях каждой точки опоры делало исследование объёмных параметров, геометрических характеристик не только повседневной работой, но и практически ежеминутной работой. В связи с этим совершенствуется бинокулярное зрение, стимулируется развитие коры на границе затылочных и теменных долей – стимулируются ассоциативные зоны коры, и, несмотря на то, что реально работает уровень С, структуры этого уровня готовят кору к работе на уровне Д – во первых, объединяются в единый образ зрительные характеристики ветвей и лиан с тактильными ощущениями от них же, и, во вторых, для сенсорных коррекций каждого шага в трехмерном пространстве закладываются связи между нижней теменной долькой и премоторной корой. Впрочем, нижней теменной дольки на этом этапе развития еще нет как таковой, речь идет о структурах-предшественниках. Тем не менее, прыгая с ветки на ветку, животное должно по зрительному образу очередной точки опоры четко представлять себе, какова она будет на ощупь, значит, функциональный аналог верхней теменной дольки не может не быть. Я не рассматриваю анатомических изменений конечностей, этот вопрос рассмотрен в сравнительно-анатомических исследованиях приматов. Следует заметить лишь, что строение конечностей при перемещении в трехмерном пространстве, и, особенно при брахиации, усложняется. Увеличивается объём движений в плечевом и тазобедренном суставе, развиваются способности пальцев на конечностях к надежному захвату опоры. Это так же требует развития управления со стороны ЦНС, более тонкой коррекции мелких мышц, что в свою очередь не может не стимулировать работу нейронов пост- и прецентральной извилин, расширение соответствующих зон становится жизненно необходимым. Здесь есть смысл сказать о том, что в физиологии есть понятие двигательных единиц. Двигательной единицей называется двигательный нейрон, и мышечные волокна, получающие от него иннервацию. Двигательные единицы подразделяются на большие и малые. В больших двигательных единицах управление одного нейрона распространяется на большое количество мышечных волокон (один нейрон может иннервировать до 20 000 мышечных волокон), в малых двигательных единицах один нейрон может управлять сокращением 1-3-х десятков мышечных волокон, а в некоторых случаях (мышцы глаза) и одно-единственное мышечное волокно. Малые двигательные единицы входят в состав дистальных частей конечностей, имеющих особое значение при перемещении по ветвям деревьев. Таким образом, сам по себе древесный образ жизни стимулировал относительное увеличение количества нейронов, ответственных за локомоции в этом образе жизни.
Таким образом, древесная жизнь обезьян заложила основы восприятия предметного уровня в структурах уровня локомоций. Разнообразие локомоторных движений в трехмерном пространстве изменило развитие ЦНС в направлении меньшего сохранения следов научения в генетическом аппарате, и большей способности к научению в постнатальном периоде. Т.е. анатомическая и функциональная незрелость центрального нервного аппарата дриопитеков с возможностями дальнейшего постнатального развития была заложена достаточно рано – на границе мезозоя и кайнозоя – порядка 70 млн. лет. Действие ФРН – фактора роста нейронов – на рост функциональных связей в нервной системе в постнатальном периоде онтогенеза в значительной степени зависит от андрогенов – мужских половых гормонов (Хазен А.М. 2000). Это значит, что половая активность кайнозойских приматов была отнюдь не сезонной, не периодической, а в достаточной степени постоянной. В этом смысле половое поведение дриопитеков мало отличалось от такового у современных человекообразных обезьян. Это существенное замечание. В дальнейшем мы увидим, что особенности полового поведения оказали сильное влияние на эволюцию приматов.
Совершенно очевидно, что ход эволюции обезьян был тесно связан с тектоническими процессами преобразования поверхности Земли. Тогда формировались мощные горные складки, климат менялся с тропического влажного на умеренный, а где-то и холодный и сухой. Обезьяны были поставлены перед выбором – погибнуть, измениться или мигрировать. Плейстоценовые формы азиатских обезьян преобразовались, дав начало современным формам обезьян и предку человека. Эти обезьяны в большинстве своем были вынуждены перейти с древесного образа жизни на наземно-древесный образ жизни, а некоторые на чисто наземный, как многие павианы. Предки человека должны были спуститься с высот на землю.
Наземный образ жизни привел к прямохождению, что эпизодически демонстрируют и современные человекообразные обезьяны. Прямохождение в свою очередь вызвало соответствующие изменения в морфофункциональных параметрах тел – удлинение детства, освобождение верхних конечностей от функции локомоции, форма позвоночника и многие другие. Причина же именно прямохождения после прекращения древесного образа жизни на сегодня не совсем понятна. Сам факт прямохождения не подлежит сомнению, но как случилось, что предок человека предпочел хождение на двух ногах, явно более трудное и медленное, чем использование для локомоций всех четырех конечностей? Для этого нужно было получить какое-то явное преимущество, перед которым локомоторные потери от прямохождения стали бы несущественными. Большинство авторов не отрицают значения для прямохождения использования верхних конечностей для манипуляций с предметами, но прямого однозначного ответа все же нет. Причина в том, что недостаточно материала для анализа и обсуждения всех условий, приведших к устойчивому прямохождению после прекращения древесного образа жизни. Нет прямого ответа и у нас. Но некоторые соображения мы считаем нужным привести.
Среди теорий есть и явно бредовые, вроде теории о водном этапе антропогенеза. В то же время в этой теории затронут важный вопрос, игнорируемый прочими теориями. Речь идет об утрате шерсти. Водный этап антропогенеза предусматривает после себя не только прямохождение, но и изменение кожного покрова. Мы же считаем, что утрата шерсти предками человека, несомненно очень важный момент, и к нему мы вернемся немного позже. Здесь есть смысл рассмотреть археологические находки иного характера и возможно они помогут немного больше сказать о факторах антропогенеза, действовавших примерно в то же время. Речь пойдет о строении зубов (рис. 6).
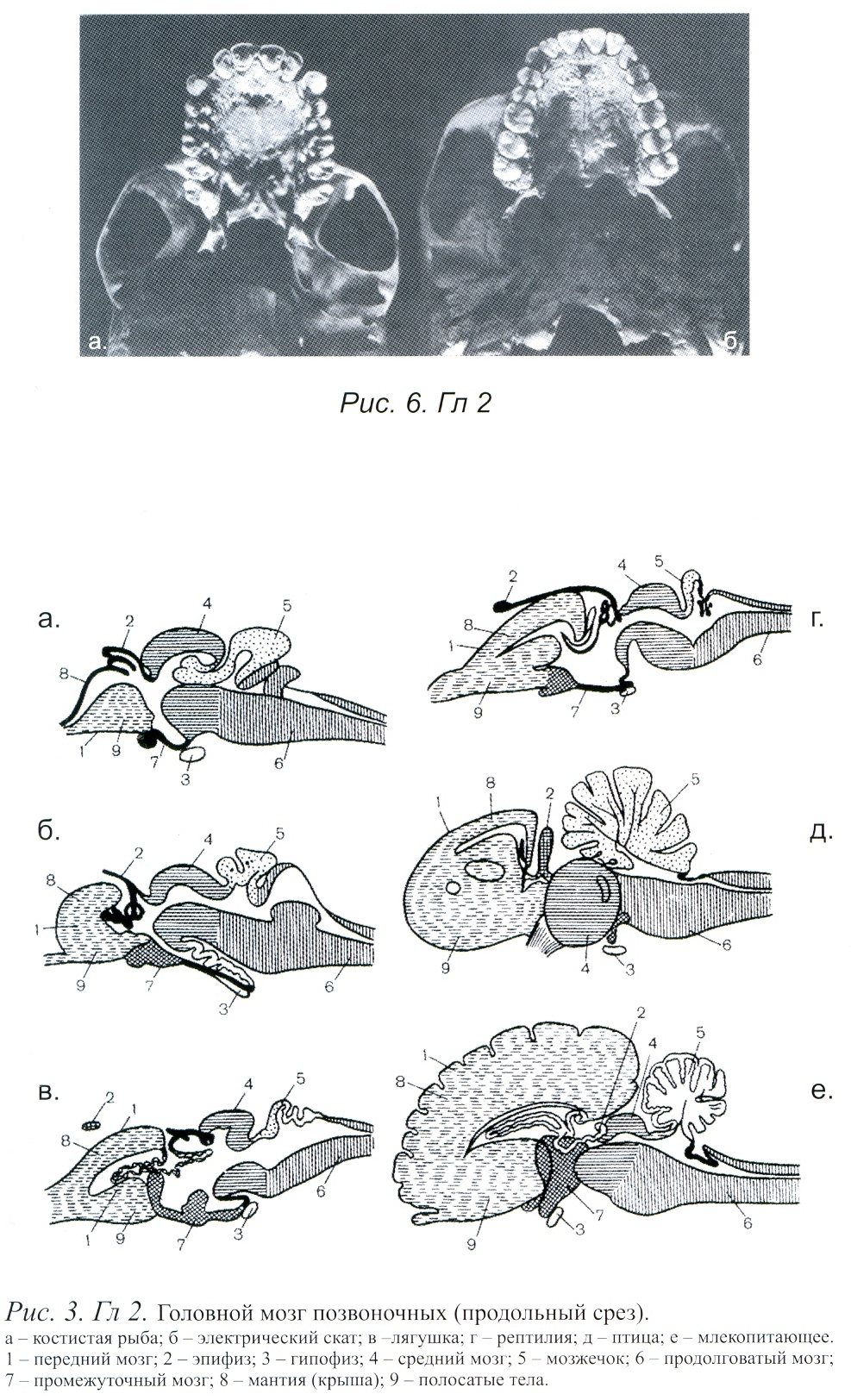
Спутать верхнюю челюсть человекообразной обезьяны (вверху) с челюстью гоминида (внизу) невозможно. В челюсти обезьяны (в данном случае — современного карликового шимпанзе) зубы располагаются по трем сторонам прямоугольника, причем ряды коренных зубов почти параллельны. Четыре резца на передней стороне отделены от других зубов промежутками, в которые входят длинные клыки, характерные для самцов всех человекообразных обезьян.
Человеческая челюсть не прямоугольна, а подковообразна. Зубы располагаются по дуге, расширяющейся в глубине рта. Поскольку у людей клыки невелики, нужды в просветах нет, и все зубы расположены тесно. У человека коренные зубы крупнее резцов, тогда как у обезьян вверх они по величине почти не различимы. Кроме того, человеческая челюсть и не выдается далеко вперед.
Различие в форме челюстной дуги дополняются различием в возможностях движений нижней челюсти относительно верхней. Височно-нижнечелюстной сустав человека устроен так, что допускает движения вверх-вниз, вправо-влево и вперед-назад. У человека нет клыков, которые мешали бы ему двигать челюсть в стороны, то есть совершать полноценные жевательные движения. Уплощенный ряд боковых зубов более приспособлен к перетиранию твердой растительной пищи, чем остроконечные моляры обезьян. Современные человекообразные обезьяны более хищники, чем человек. Возражение о том, что человек приобрел такие зубы недавно, отметается. Аналогичные зубы с остатками челюстей найдены на территории южной, центральной Азии и Европы, и датируются как произведения 5-6 млн – летней давности. Давний предок человека, намного более давний, чем австралопитек, питался на протяжении миллионов лет грубой растительной пищей, скорее злаками. Или скажем так – значительное время в году рамапитеки вынуждены были питаться зерно-бобовыми и корнеплодами. Исходя из предполагаемого потенциала, позволившего дальнейшее развитие, этот рацион мог быть дополнен белковой пищей – насекомыми, мелкими животными, поедание которых не оказывало большого влияния на зубочелюстной аппарат, а значит, было эпизодическим; и по сезону – фруктами. В теплых тропиках и субтропиках подобный режим питания на протяжении миллионов лет был бы невозможен. Изобильная флора и фауна не создает условий для пристального внимания к зерно – бобовым, здесь слишком много плодов, фруктов достаточно мягких по консистенции. Животная пища, все же требует развития клыков и режущих моляров, что мы и видим у человекообразных обезьян.
В плане экологическом, он конкурировал со всем известными грызунами – мышами и крысами. Да, не найдено никаких следов строительной деятельности такой давности. Но подобная деятельность не в диковинку в живой природе. И уж если крысы и мыши могут себе устроить жилище, то нашему предку сам бог велел этим заниматься. Подобная деятельность, с точки зрения физиологии движения не является предметной – это врожденное свойство, закрепленное в памяти хромосом определенных видов. Грызуны могут миллионами лет рыть норы, от этого уровень Д у них не продвинется ни на шаг. И рамапитек был полноценным животным. В отличие от мышей он прошел древесную стадию жизни, уровень его локомоций уже включал в себя элементы предметной деятельности в структурах мозга, ответственных за локомоции. Мы ничего не можем сказать о жилище рамапитека. Но его не могло не быть. В каком виде? В виде нор, гнезд, шалашей, каменных завалов, покрытых сверху бревнами. Если мы не предположим наличие жилища у рамапитеков, жизни в колониях, подобно многим грызунам, нам нечем будет объяснить параллельное развитие следующих признаков: развитие уровня Д, утрата шерсти, возможности длительного обучения детенышей сложному виду локомоций – прямохождению у потомков рамапитеков – австралопитеков, развитие социальных основ жизни современных человекообразных обезьян.
Оседлый образ жизни рамапитеков, а скорее всего, не всех, а живущих на северной, самой холодной границе ареала, возможен. Он не противоречит ни одной археологической находке, хотя и не подтверждается, но скорее всего, и не может быть подтвержден – слишком много прошло времени, остатков просто не найти. Да и искать их надо не в южных теплых краях. Необходимо напомнить, что основные находки австралопитеков сделаны в Африке, рамапитеков в Азии и восточной Европе, большей частью в тех местах, где климат в наше время теплый. Холод же играл, и по прежнему играет важнейшую роль в антропогенезе, но места, где холодно сейчас антропологов мало привлекают.
Древесные обезьяны уже получили в оформлении структур уровня С (стрио-паллидарная система, первичная кора) возможности уровня Д, принципиально не отличались по строению ЦНС от прочих млекопитающих, то с возникновением прямохождения, в ЦНС стали происходить принципиальные изменения. Эти изменения Бернштейн называет развитием уровня предметных действий (Д). Освобождение передних (верхних) конечностей от функции локомоций сделало ненужным нервный аппарат, управляющий этими конечностями как органами перемещения. Но поскольку в этом же аппарате уже были заложены возможности нового уровня, и именно эти возможности стали актуальны для свободных верхних конечностей, то аппарат управления стал действовать игнорируя старые возможности и используя новые. Действительно, у современных млекопитающих (по Бернштейну) предыдущий уровень управления движениями развит значительно лучше, чем у людей. Животные быстрее, сильнее, выносливее, точнее наносят удары, во всех отношениях их локомоции совершеннее. Но уровень предметных действий, в принципе доступный многим млекопитающим, ни в коей мере не сравним с таковым у человека. Таким образом, именно развитие уровня предметных действий сочеталось с утратой преимуществ четвероногости. Именно качества локомоций предок променял на предметные действия. В какой ситуации это возможно? Только в одной – оседлый образ жизни.
Оседлый образ жизни предполагает наличие убежища. Тогда есть возможность без большого ущерба для безопасной жизни утратить возможность эффективной локомоции. Достаточно вовремя заметить опасность и сделать один мощный рывок до убежища. При этом требования к организму по выносливости снижаются, зато повышаются требования по спринтерским качествам. Спринтерские же качества, есть качества, опять же, не столько опорно-двигательного аппарата, сколько, прежде всего, качества нервной системы. Их два: первое – по Павлову – преобладание в обмене веществ мозга над печенью. На уровне обмена веществ – преобладание серо-содержащих аминокислот над азот-содержащими аминокислотами. Расширение спектра веществ, регулирующих метаболизм. Второе – организация общественных отношений. Просто необходимо создать общественную службу оповещения об опасности. Любое разделение труда возможно только при непрерывно действующих общественных отношениях. Общественные же отношения требуют постоянной активности тех частей мозга, которые ответственны за взаимоотношения – лимбическая система (эмоции) и влияние нижних отделов прецентральной извилины (мимическая мускулатура). Если к этому добавить появление и развитие третичной коры, в теменной доле, а так же внимание к звуковым сигналам наблюдателей, то мы получаем то что надо. Это было именно в самой холодной части ареала – утепление убежища предполагает непрерывное совершенствование предметных действий. Ничего необычного в этом нет — примеры строительства жилья животными любой присутствующий приведет без труда. Современные обезьяны так же строят себе гнезда или шалаши, но не живут в них долго. У них нет для того причин – в Африке тепло.
Есть ли аналог сегодня в природе той ситуации? Да есть. Грызуны. Могут ли грызуны стать на место человека – нет. Им не хватает полноценного бинокулярного зрения – не тот онтогенез. Отсюда образ предмета не может захватить внимание грызуна с той силой как у нашего предка.
Второй вопрос – что заставило жить в домах? Только одно – похолодание, от которого невозможно откочевать на юг. Именно похолодание стало фактором, исчезновения тропических лесов, согнавшим наших предков на землю, но оно же и заставило их строить дома. Те, у кого была возможность откочевать на юг – остались обезьянами.
Но жизнь на островке жизни, сопровождалась еще и сменой питания. Холод наверняка уничтожил теплолюбивые растения, пришлось питаться тем, что осталось. Смена типа питания привела к нестабильности генома, каковая, давая множество мутаций, позволила нашим предкам измениться – в мозгу произошли изменения, необходимые для совершенствования предметных действий, появилась третичная кора, усилилась лобная кора. В поведении усилился общественный компонент.
Таким образом, наш предок ходил на двух ногах. Был он покрыт шерстью. Дальнейшее развитие пошло по пути приспособления к холоду. Я сознательно не касаюсь конкретного момента времени. Заостряю внимание на главном факторе естественного отбора – похолодание.
Дело в том, что похолодание вносит свой, вполне определенный вклад в изменение флоры и фауны. В поисках замены утраченным, привычным источникам пищи, животные не могут не перейти на новые источники питания. Меньше растительной пищи – больше внимания к пище животной и наоборот. Таким образом, эти изменения приводят к смене питания наших предков, а смена типа питания становится мощным мутагенным фактором.
Необходимо добавить, что, встав на путь предметных действий, рамапитек совершал их строго по крайней необходимости. Когда были исчерпаны все возможности уровня С, животное может, при наличии необходимых структур в ЦНС совершить нечто подобное. Но это не уровень Д — это именно уровень локомоций, усовершенствованный особо сложными локомоциями, и закрепившийся в геноме. Как умение бобров строить сложные гидротехнические сооружения.
В условиях же нарастающего холода возникает необходимость не только строить жилье, но и заботиться о сохранении в нем тепла. Заботясь о потомстве, спасая детенышей от холода, пришлось создать подобие одежды для них. Жилье и одежда сформировали микроклимат вокруг тела. Этот фактор быстро привел к утрате шерстяного покрова. Так предметная деятельность стала причиной закрепления случайных изменений в геноме.
Кроме того, в Африке пища в изобилии круглый год. Обезьяны перемещаются с места на место в поисках пищи. Место жительства нашего предка было таким, что холод там все усиливался, а значит холодный период – зима – все удлинялся. Возникла необходимость совершать еще один вид предметной деятельности – запасать пищу на зиму. И в этом ничего необычного среди животных нет – многие грызуны это делают. Но запасы невозможно переносить при миграциях. Зимой надо жить там, где твои запасы. Вот что привязывало предков людей к месту.
Когда произошла утрата шерсти? Археологические находки, сделанные в поисках предков людей ничего не могут сказать о наличии или отсутствии шерсти у этих предков – находят только кости и зубы, шерсть не может храниться в земле даже несколько лет. Но мы можем с большой степенью уверенности предполагать, что укротители огня имели голую кожу. Дело в том, что животному, покрытому шерстью близость огня смертельно опасна. Шерсть сама по себе является теплоизоляцией – это ее основная функция у теплокровных. В природе огонь проявляет себя как стихия – внезапно возникает и внезапно исчезает. Даже замерзшее животное, нуждающееся в тепле, если при внезапно возникшем лесном пожаре попробует погреться у огонька рискует поджечь шерсть. Сухая шерсть достаточно хорошо горит, но если даже её, горящую, потушить, могут остаться ожоги. Ожоги плохо лечатся даже в человеческом обществе, ожог третьей степени (с повреждением мягких тканей) труднее вылечить чем ссадину той же площади. Страх животных перед огнем не носит никакого мистического характера. Это закрепленная в хромосомном аппарате реакция избегания опасности. Миллионы лет жизни в домах, в колониях, когда младенец защищен от естественного климата данной местности стенами помещения, а в некоторых случаях вокруг его тела создан микроклимат в виде сена, пуха, перьев, которыми заботливая мамаша защищала дитя от холода, привели к тому, что шерсть была утрачена. Мамаша же совершенствовалась в предметной деятельности.
У неё постоянная повседневная потребность в новой и новой предметной деятельности была обеспечена. Это утепление жилья. Утепление младенца – первый шаг на пути создания одежды. Создание запасов пищи на зиму. После этого, наш утративший шерсть предок не мог не стать человеком, но еще не был им. Защищаясь от холода, наш предок имел постоянную работу для рук и для мозга. Он обрел тренированный в предметной деятельности мозг и утратил шерсть. Еще раз – неизвестно, когда была утрачена шерсть, но известно, что первые следы пользования огнем найдены в Африке и датируются около 800 000 – 700 000 лет назад.
Таким образом, период около 10 млн лет назад продолжительностью несколько миллионов лет был потрачен рамапитеком на обучение прямохождению. Развитие этого качества развивалось медленно, в соответствии с меняющимися природными условиями. Возможно, было несколько параллельных видов, обретших прямохождение, возможно, какие-то из них утратили это свойство. Установить точное продвижение по эволюционной лестнице не представляется возможным. Важно, что при этом появившиеся возможности были реализованы в направлении развития коры головного мозга и в первую очередь ассоциативных зон, позволивших перейти от локомоций к предметным действиям. Ведущим мотивом на уровне действий становился не предмет сам по себе, а смысловая сторона действия с предметом – как объектом или орудием. Возникали и закреплялись в хромосомном аппарате связи между отражениями предмета (ассоциативная кора между затылочной и теменной доли формирует понятие предмета) и смысловой структурой действия (ассоциативная кора между затылочной, теменной и височной долями), и формированием акцептора результатов цепи последовательных действий (премоторная кора).
Появление первых каменных орудий относится ко второму миллионолетию до нашей эры. Это значит, что 8 миллионов лет прошли не зря, умение работать с предметами, оставившее следы на камне прошли не зря. Австралопитеки ходили на двух ногах, обрабатывали с таинственной целью камни. Магистральная линия антропогенеза подразумевала постоянную необходимость совершенствования предметной деятельности. Эта деятельность была до предела прагматична. Не баловались тысячелетиями австралопитеки. Они использовали каменные орудия, а вместе с ними и деревянные, не способные к длительному хранению для того, чтобы жить самим, и давать жить своим потомкам. Понятно, что не все пути вели к человеку. Там где жить было легко, по крайней мере, с накопленным эволюционным багажом оказаться в более благоприятных условиях, там не было нужды напрягаться, использовать все резервы организма, и его управляющего органа, там была возможна деградация, там располагался тупик эволюции. Это были вполне человекообразные животные, способные к изготовлению каменных орудий. Но эти орудия были им нужны для жизни. Научившись изготовлять и использовать орудия в тяжелых условиях, никто не хотел их совершенствовать, когда условия становились легкими. Тем не менее, около 0,8 млн лет назад австралопитеки получили возможность пользоваться огнем. Значит до этого был длительный период пользования одеждой, по крайней мере для утепления младенцев. Младенцы утратили способность выращивать на себе теплую шерстяную шкуру. При этом зубы сохранили следы поедания зернобобовых, при этом австралопитеки охотились в том числе и на крупную дичь – вплоть до слонов. Единообразие каменных орудий говорит о том, что в изготовлении их прогресса не было, то есть их изготовление – не собственно предметная деятельность в современном человеческом понимании, это закрепленная в хромосомах автоматизированная деятельность, в овладении которой все большее значение имела педагогическая деятельность старшего поколения и все меньше – врожденная склонность манипулировать предметами. Прожив несколько тысяч лет закутанными в шкуры, младенцы утратили собственную шерсть. Это означает, что прямая линия антропогенеза подразумевала к тому времени жизнь в жилище, длинное детство в нем, зачатки одежд, запасы пищи на трудный сезон – короче – оседлый образ жизни. Кочевая жизнь либо была вынужденной и временной, либо означала уход в сторону от генеральной линии антропогенеза.
И вот тут то эволюционное развитие двинулось вперед семимильными шагами. И тому несколько причин.
1. Голая кожа не защищала от холода, и предметная деятельность стала вопросом выживания. На всю жизнь появилась потребность делать одежду. Жилье и одежда стали необходимостью и их утепление стимулом к форсированному развитию предметной деятельнсти.
2. Увеличение рецепторного поля за счет термочувствительности привело к стимуляции именно теменной доли. В образ предмета свой вклад стала вносить термочувствительность, и он обогатился и расширился. Глаз стал различать не только степень опасности огня для жизни, но и достаточно точно определять температуру огня. Последовательность превращения предметов при термообработке, необходимость отслеживать в течение определенного времени температуру таких предметов (ждать, когда остынет), включилось в смысловую структуру действия.
Освоение огня стало еще одним глобальным фактором, стимулирующим предметную деятельность:
1. Революцию в эволюции произвела термообработка пищи. Термическая обработка делала пищу более энергонасыщенной и, кроме того, облегчала все этапы пищеварения. Уменьшение нагрузки на жевательный аппарат делало его избыточно эффективным для жевания, следствием чего становилась гипотрофия и частичная атрофия его анатомических составляющих. Все это привело к формированию морфофункциональных предпосылок к созданию речи – облегчение зубочелюстного аппарата, повышение точности его движений.
2. Огонь, как фактор влияния на хромосомы. Речь в данном случае идет о двух моментах. Во-первых, при горении образуются непредельные полициклические углеводороды, которые, попадая в организм с воздухом, сами по себе могут быть мощным мутагенным фактором. Во-вторых, обожженная пища содержала в себе еще больше таких продуктов.
После огненной революции эволюция продолжается в новых условиях. Мы получаем ситуацию – австралопитек, прямоходящий, строящий дома, одежду, делающий запасы на зиму, с голой кожей и освоивший огонь.
Возникли анатомо-функциональные предпосылки для формирования второй сигнальной системы. Это 1. развитый уровень Д., 2. уменьшение пищевой нагрузки на жевательный аппарат. 3 общественный образ жизни.
Но надо понимать, что это не люди. Они многое могут из уровня Д, но большая часть их умений есть результат не столько педагогической передачи знаний от поколения к поколению, а результат закрепления в хромосомах умения совершать действия, подобно бобрам. Поэтому педагогам — австралопитекам нечему учить учеников, те уже только родившись, многое знают. В нужный момент они захотят ходить на двух ногах, в свое время построят жилье (когда придет пора размножаться), к одежде приучит мать, и только работа с огнем, в силу того, что очень мощные структуры в организме созданы для обеспечения страха пламени потребует специального обучения. И снова и снова мамы будут кутать теперь уже навсегда потерявших способность отращивать собственную шерсть детенышей, при наступлении холодов, а папы – совершенствовать утепление жилища. А там где холода не наступают – там возникают боковые ветви антропогенеза.
И так будет продолжаться еще полмиллиона лет, пока на сцене не появится тот, кого мы называем неандертальцем.
2.4 НЕАНДЕРТАЛЕЦ И КРОМАНЬОНЕЦ
Примерно 100 тысяч лет назад на земле появился неандерталец.
Его предки уже давно пользовались огнем, не имели шерсти на коже, и не менее двух миллионов лет ходили на двух ногах, могли жить в домах и носили подобие одежды. Говорить они не умели. Каменные предметы, изготовленные австралопитеками, за два миллиона лет изменились мало, то есть изготовление каменных орудий каждым поколением изготовлялось практически стереотипно. Не было серьезных попыток усовершенствовать эти орудия, эволюция шла по обычному пути, головной мозг менялся в филогенезе в направлении цефализации, но выйти на уровень Д, в полном объеме, так, чтобы включение уровня происходило не в крайнем случае, а сделать его работу обычной повседневной практикой у австралопитеков не получилось. Тем не менее, уход за младенцами требовал использования материалов для укрытия от холода. Наилучшим таким материалом были, несомненно, шкуры животных. Скорее всего, именно манипуляции с сырьем кожевенной промышленности занимали большую часть предметных действий австралопитека. Снять шкуру с животного, аккуратно очистить ее от жира, обработать ее дубильными веществами, размять, чтобы была мягче – вот основные этапы работы с кожей.
 Именно для этих целей были наиболее приспособлены большая часть каменных изделий тех тысячелетий. Дубить шкуры не составляло большой сложности – любой младенец сделает это легко, и не один раз за сутки (моча обладает дубильнымм свойствами). Из чего следует, что охота составляла обязательную часть претендентов на звание царя природы. А главным стимулом к развитию умственных способностей особей женского пола была забота о младенцах – все более беспомощных при рождении.
Именно для этих целей были наиболее приспособлены большая часть каменных изделий тех тысячелетий. Дубить шкуры не составляло большой сложности – любой младенец сделает это легко, и не один раз за сутки (моча обладает дубильнымм свойствами). Из чего следует, что охота составляла обязательную часть претендентов на звание царя природы. А главным стимулом к развитию умственных способностей особей женского пола была забота о младенцах – все более беспомощных при рождении.
Уже у поздних австралопитеков уход за новорожденными требовал от матери не только внимания и труда, но и знаний, которые могли быть получены только от старшего поколения, от более опытных товарищей, которые сами имели детей, сами имели опыт ухода за ними, и сами получали дополнительные знания от старших товарищей. Именно в этом проявлялась культура (знания, передаваемые по наследству последующим поколениям), а не в способе обработки камня, каковой почти не менялся на протяжении миллионов лет. Уход же за детьми требовал использования не только высших на тот момент технологий, но и вследствие эволюционных изменений в теле животного, усовершенствования этих технологий. По крайней мере, именно у неандертальцев мы увидим четкие следы того, что уровень предметных действий стал практикой повседневности.
 первобытные люди обрабатывают шкуры животных
первобытные люди обрабатывают шкуры животных
На этой диаграмме (рис. 7) прослежен постоянный прогресс в способах изготовления кремневых орудий. Она показывает, как увеличивающееся число ударов (кружки) и разных операций (группы кружков) вели к появлению лучших орудий и к более продуктивному использованию сырья. Наиболее примитивное орудие требовало 25 ударов и одной операции, тогда как самое позднее и самое отделанное получалось после 251 удара, разделенных на девять сложных операций, которые требовали от каменных дел мастера высокой точности, глазомера и сообразительности. (последняя технология будет показана позже).
 рис. 7
рис. 7
Появление неандертальцев и их исчезновение прошло на протяжении примерно 60 000 лет. Какой же путь они прошли, передав в конце своего пути эстафету кроманьонцам? Казалось бы, австралопитеки сделали большую часть работы. Дом, одежда, огонь – немало. Но именно неандертальцы сделали уровень Д постоянно действующим. Им не удалось довести уровень Д до нашего современного уровня, но сделанное ими подвело эволюционную линию вплотную не только к возможности развития уровня Е (речь), но и сделали этот переход жизненно важным для кроманьонцев, обеспечили неизбежность такого шага.
Что мы знаем о неандертальцах. Появились на земле примерно 100 000 лет назад и исчезли 40 000 лет назад. Они распространились по Азии, Африке и Европе. Их каменные орудия были более совершенны, чем орудия австралопитеков. Использование огня, скорее всего, неандертальцы использовали на всем протяжении отпущенного им срока земного бытия. Они вели и оседлый и кочевой образ жизни. Следовательно, они могли жить и в постоянном жилище, и во временных переносимых домах. Мужчины могли заниматься охотой, мясной рацион был достаточно обычным, термообработка пищи тоже не была экзотикой.
Общественное устройство у неандертальцев во всех подробностях нам неизвестно. Но некоторые твердо установленные факты говорят о том, что элементы системы социального обеспечения у неандертальцев были в ходу. Искалеченные и состарившиеся члены групп неандертальцев оставили свои скелеты со следами тяжелых травм с последующим неполным восстановлением и тяжелых хронических болезней с длительным течением. Например, мужчина из Ла-Шапель-о-Сен был буквально скрючен артритом, и не мог участвовать в охоте. Ел он с трудом, так как давно потерял все зубы, кроме двух. Возможно, соплеменники помогали ему жевать пищу. Некоторые находки говорят о том, что люди с врожденными дефектами вырастали, становились взрослыми и умирали в преклонном возрасте, явно не будучи ни полноценными охотниками, ни работниками физического труда.
Таким образом, мы подходим к очень важному моменту в жизни неандертальцев. Общество неандертальцев было именно обществом. Слабые и больные в нем могли рассчитывать на помощь со стороны соплеменников, на кусок пищи, место у огня, защиту от хищников. Трудно сказать, насколько распространенными были подобные вещи. Может быть, такое было не во всех племенах и не всегда, мы не знаем, на какие категории лиц распространялась такая помощь. Важно, что в обществе неандертальцев было то, что заметно отличало его от сообществ животных. У животных в природе можно встретить нечто подобное, но не в таких масштабах, как у наших предков.
Дело в том, что длительная забота о младенцах, ее постоянное совершенствование и, возможно, передача каких-то умений в этой области от поколения к поколению создали определенное отношение у женщин, видимо, в первую очередь, к слабым и немощным. Забота о младенцах требовала такой квалификации, и, когда младенцы становились независимы от матери, материнское умение и стремление опекать не находило себе применения, но требовало своей реализации. Умения, доведенные до автоматизма, становятся навыками, а навыки могут проявиться без участия сознания при возникновении ситуации, в которой они были обретены. Женщины-неандертальцы должны были уметь многое – утеплять жилище, обрабатывать шкуры зверей, делать одежду, поддерживать огонь, готовить на огне пищу, принимать роды, ухаживать за младенцами, и видимо, за больными и увечными. Все это им обеспечивал уже развитый уровень Д. Напоминаю, что о речи не было и речи. Часть навыков были врожденными, но значительная часть уже приобреталась на протяжении жизни. За 60 000 лет количество врожденных навыков уменьшилось, а на долю приобретенных приходилось все больше и больше. Охота мужчин и собирательство женщин тоже совершенствовались, иначе откуда взяться прибавочному продукту для поддержания жизни нетрудоспособных. Но совершенствование навыков при этих занятиях было вторичным, следствием уже развитых отделов мозга, необходимых для обеспечения продолжения жизни. Навыки могут быть врожденные и приобретенные. Для развития врожденных навыков необходим отбор особей, случайно способных совершать нужные для жизни действия, и их закрепление в хромосомах происходит на границах ареалов. Приобретенные навыки, то, что Павлов назвал условным рефлексом, тоже может приобретаться при случайно возникших обстоятельствах. Но целенаправленному обретению навыков может способствовать общественная среда. У человека мы это видим при обучении детей от родителей и в школе.
У неандертальцев общество на протяжении 60 000 лет совершенствовалось в целенаправленной передаче навыков из поколения в поколение посредством воспитательного процесса. Каждая мать, становясь бабушкой, обучала свою дочь приемам определенных очень сложных действий. Эти приемы воспринимались дочерью как умения, то есть требовали от нее на этапе обучения максимального внимания, максимального тонуса коры головного мозга. По мере повторения действий и превращения умения в навык, новейшая кора головного мозга, обучив старую кору или подкорковые центры, участвовала минимально в обеспечении действия. Со стороны матери этот процесс не мог быть автоматическим. Каждая новая дочь отличалась от предыдущей по способностям к тем или иным действиям, как мы это видим и сейчас у наших детей. Обучение второй дочери отличалось в деталях от обучения первой, и требовало от матери новых ментальных усилий. Показ своим примером действий снова и снова носил характер символический. Таким образом, символические действия были повседневной практикой женщин неандертальцев. О речи говорить еще рано. Все это делается молча, сопровождаясь эмоциональными звуками. Одобрение и неодобрение – все это жестами, мимикой, мычанием (как коровы).
Обучение требовало обязательного созревания структур организма, необходимых для выполнения действия. Мужчины и женщины специализировались в разной деятельности. Поскольку младенцы росли и развивались все медленнее, женщины, начиная со второй половины беременности и до конца вскармливания грудью, были беспомощны за пределами общества. Особенно это касается мест, где климат не позволял обходиться без жилья и одежды. Мы полагаем, что в теплом климате нет смысла искать предков человека. Теплый климат и большая биологическая продуктивность местности исключают необходимость постоянной заботы о тепле и пище, а, стало быть, и необходимость в развитии коры головного мозга.
Женщины были ответственны за передачу знаний и умений по женской линии – в первую очередь связанных с сохранением и выращиванием детей, что впоследствии привело естественным образом к заботе о больных и травмированных. Для женщин было важно передать свои умения, у них и совершенствовалась коммуникативная сторона. Умение передать умение и есть знание. Вот мы и добрались почти до человеческого в неандертальцах.
Для мужчин важно было, вступив в возраст половой зрелости обеспечить женщин возможностями жить нормальной женской жизнью. В то же время, вырастая рядом с мамой, глядя на мир мамиными глазами, мужчина должен был оторваться от нее. Умение быть мужчиной в большинстве племен означает умение быть охотником. Охота же неандертальцев, в принципе не отличается от охоты других животных. Использование оружия и огня были однократными прорывами мужчин в уровень Д. Большая часть действий на охоте вполне обеспечена уровнем локомоций, уровнем С. Животным, ведущим групповую охоту вполне достаточно тех средств коммуникации, которые у них есть. Без всяких долгих или кратких разговоров, волки так могут организовать свои действия, что олень или лось будет загнан в болото, где его легко будет взять. Львы тоже охотятся группой, и тоже обходятся без речи. Мужчин людьми делало участие в воспроизводстве потомства. Я имею в виду обеспечение женщин безопасностью, пищей и жилищем.
Будучи детьми своих матерей, мальчики вполне могли научиться именно их умению передавать умение. На охоте они просто подражали старшим, это подражание не было шагом вперед, как не было шагом вперед со стороны старших и просто охота в присутствии младших.
Демонстрационный показ со стороны старших действий, которым сам научился – вот шаг вперед, по сравнению с животным миром. Демонстрационный показ для наблюдателя с целью воздействовать на этого наблюдателя, вызвать с его стороны определенные действия, причем действия не врожденные, а такие, каких наблюдатель до того не делал и, возможно не умел, и врожденной предрасположенности к которым не имел, и последствий которых наблюдатель, возможно, себе не представляет, но сделать так, чтобы у наблюдателя не возникло страха перед возможными последствиями, как, может быть, страх у птенцов перед первым вылетом из гнезда – это и есть символическое действие. (Пример с птенцами ограничен тем, что у птенцов перед первым полетом есть врожденное умение летать, как и у их родителей есть врожденное умение показывать полет).
Символическое действие означает предельное для доречевой стадии развитие уровня Д. Это означает, так же, свободное владение включением этого уровня – не в экстремальной ситуации, а в повседневной жизни. Это значит, что для неандертальцев время стало не только ритмом, не только его течение от начала к концу, они заметили его в ситуации до и после. Иначе говоря, закон причинности перестал быть тайной для неандертальцев. Что касается охоты, то ее действие на работу мозга ограничивается повышением тонуса ЦНС, с минимальным участием уровня Д, хоть с палками, хоть с ружьем.
2.5 МАГИЯ
2.5.1 Магия – свойство лимбической системы.
Среди прочих следов жизнедеятельности неандертальцев самыми неожиданными были признаки их ритуальной деятельности. В 1938 году в Узбекистане в пещере Тешик-Таш было обнаружено захоронение неандертальского мальчика. Могила сохранила признаки погребального обряда – шесть пар козлиных рогов были вкопаны в землю вокруг могилы, образуя подобие кольца. На севере Ирака в 1960 году в пещере Шанидар, в слое давностью 60 000 лет была найдена могила неандертальца. Образцы грунта, взятые из могилы, были исследованы в лаборатории французского Музея Человека. Палеоботаники обнаружили в образцах огромное количество пыльцы, совершенно невероятное для случайного ее скопления. Ни птицы, ни звери, ни ветер, не могли занести столько цветов в глубину пещеры и положить их под могильные камни. Несомненно, покойный был буквально засыпан цветами. Многие другие находки говорят о том, что неандертальцы серьёзно относились к умершим. Они совершали над ними обрядовые действия. Зная, что закон причинности был знаком неандертальцам, по крайней мере на уровне пользователей, мы не должны удивляться тому, что они вполне могли совершать символические действия – это действия развитого уровня предметных действий (Д). Закон причинности гласит – у всякого события есть причина, у всякого действия есть последствия. Следствием закона является понимание, что если понятна причина смерти, то после смерти так же должны быть последствия. Что там происходит с покойником за порогом жизни и смерти неизвестно, но что-то наверняка происходит, и это происходящее с покойником вполне может иметь какие-то последствия для живущих.
Основания для такого заключения у неандертальцев, конечно же, были – ведь не все события в их жизни имели видимую причину. Все, что не имело причины, вполне могло быть последствием действий умерших. Неплохо бы заручиться их доброжелательством, раз уж не смогли уберечь их от смерти, и, тем более, умершие не без вашего участия требовали какого то особого отношения после смерти. Несомненно, магия есть естественное следствие закона причинности и ограниченности нашего знания.
Кроме интеллектуальных основ для магии есть и иные основания.
Сколько у нас сигнальных систем?
Павлов ввел понятие вторая сигнальная система. Первая сигнальная система – собственно органы чувств и отделы нервной системы, обеспечивающие их работу. Вторая сигнальная система – речь. В то же время, очевидно, что животные великолепно решают вопросы совместного действия, не имея второй сигнальной системы. Наблюдая за животными, мы можем легко увидеть, что они достаточно легко передают друг другу информацию о собственном состоянии. По крайней мере, эмоции млекопитающих достаточно легко читаются.
Из повседневной жизни мы знаем, что эмоции и эмоциональные слова используются для воздействия на эмоции же окружающих. Информация, передаваемая выражением эмоций, влияет на эмоции же. Это более древняя информационная система. Вторая сигнальная система (по Павлову) – более молодая, принадлежит человеку.
Есть смысл выделить не две, а три сигнальные системы. Первая сигнальная система (по Павлову) – органы чувств. Эта система создает информационный поток непосредственно от природы и ее проявлений. Эта система избирательна по степени значимости для жизни явлений-сигналов. В этой системе происходит взаимодействие субъекта и объектов.
Второй сигнальной системой (в порядке возникновения в филогенезе) правильнее назвать язык эмоций. Эта система создает информационный поток, идущий только от живых существ. Она избирательна по степени важности живого существа на данный момент для данных обстоятельств. Эта система, в отличие от первой, подразумевает сильное взаимное влияние двух субъектов. Каждый участник взаимодействия в этой системе является одновременно и субъектом и объектом, учителем и учеником, управителем и управляемым. Межвидовое эмоциональное общение происходит в соответствии с природой участников, мало и редко изменяется в онтогенезе и в контексте данной работы не рассматривается. Внутривидовое эмоциональное общение может совершенствоваться на протяжении жизни. Это основа возникновения животных сообществ. Демонстрация личной силы имеет первостепенную важность при использовании этой системы.
Об органах этой системы и их развитии. Именно эмоции выражаются действием тела. В движениях проявляется субъект. Восприятие чужих эмоций и правильное их истолкование имеет жизненно важное значение при половом размножении. Поэтому именно во взаимоотношениях между особями разного пола природой отслеживается правильное прочтение эмоций. Самцы и самки не имеют права ошибаться при воспроизводстве потомства. Готовность партнера к совокуплению на разных стадиях, переход готовности из одной стадии в другую, правильное реагирование на знаки, подаваемые партнером на каждой стадии – все это важно, все это требует внимания к поведению партнера с одной стороны; и умение проявить для партнера свою готовность или неготовность к соответствующим данному моменту отношений действиям, перевод с языка собственного гомеостаза на язык внешне видимых действий, поз, танца, жестов, звуков с другой стороны, все это должно быть оформлено в виде программы действий с использованием наличного анатомически оформленного аппарата. Сюда входят как внешнее строение тела, подразумевающее его внутреннее содержание, и закрепленные в строении ЦНС центральные аппараты функциональных систем. У высокоразвитых организмов эти системы требуют в значительной степени обучения соответствующему поведению, у низкоорганизованных организмов обучение половому поведению может и не требоваться. Чем выше животное стоит на эволюционной лестнице, тем больше оно способно к обучению, и чем больше животное должно обучаться каким то действиям после рождения, тем меньше у него имеется готовых, данных к моменту рождения программ действия. Создание программ по мере возникновения надобности в них позволяет не забивать геном лишней информацией для передачи потомкам, у высокоорганизованных животных этим занимается не геном, а родители. По сравнению с первыми организмами значение генома у человека уходит на второй план. Жить в обществе могут и те, кто в природе просто не сумел бы жить.
Грубо говоря, функция памяти вида переходит от хромосомного аппарата к ЦНС, что мы и видим в наибольшей степени у человека. Это обеспечивает высокоорганизованному животному способность находить наиболее точный ответ на вызов жизни, причем именно на этот вызов, в точно тех наличных условиях, которые невозможно учесть при работе только хромосомной памяти.
Таким образом, мы видим, что если поведение между особями разного пола для гарантированного обеспечения размножения требует врожденных реакций (безусловных рефлексов), то для успешного проведения этой процедуры в конкретных условиях необходима определенная гибкость поведения. Конкретные же условия требуют, чтобы совместные действия были абсолютно согласованы. Согласование происходит путем демонстрирования для партнера состояния своего организма. Для этого необходимо иметь средства коммуникации. Каждый должен иметь средства для внешнего проявления своего внутреннего состояния, для посылания сигналов партнеру, и средства для считывания сигналов, посылаемых партнером. То есть, по внешним проявлениям: движениям тела, принимаемой позе, движениям конечностей, мимических мышц, издаваемым звукам определять внутреннее состояние организма партнера, степень его готовности к спариванию, переход из одной фазы к другой и т.д.
Таким образом, половое поведение особей противоположного пола при общении друг с другом есть процесс передачи информации о состоянии внутренней среды организма партнеру. Но есть и второй момент, тесно связанный с первым. Это выращивание потомства. В выращивании детенышей всегда (за редкими исключениями) принимает участие мать, и, достаточно часто отец. У разных видов длительность этого процесса в значительной степени варьирует (у млекопитающих от нескольких месяцев до нескольких лет), но, в любом случае, это процесс достаточно длительный. Родители, чаще мать, на протяжении этого периода общаются с младенцами. При этом поведение младенцев говорит матери о состоянии их организма, а мать на эти сигналы реагирует с определенной степенью адекватности. Чем адекватней, тем больше у младенцев шансов выжить. Но и младенцы, со своей стороны, в процессе общения с матерью научаются передавать ей сообщение о состоянии своего организма, и совершенствуются во все более адекватной и точной передаче этой информации. Средства передачи совершенствуются как в филогенезе, так и в онтогенезе. Анатомо-функциональные признаки этих средств мы видим буквально на лице друг у друга. Эмоции, выражаемые средствами мимики и жестикуляции и есть выражение состояния нашего гомеостаза. Подробное исследование эмоций выходит за рамки нашей работы .
Управление же эмоциями происходит определенными структурами головного мозга. Эти структуры у высших позвоночных осуществляют проявление эмоций как сигналов, выраженных в позе животного, жестикуляции, мимике и звуках. Афферентный синтез формирует картину состояния организма с учетом его внутреннего состояния и его личностного отношения к этому внутреннему состоянию и не менее личностного отношения к окружающей обстановке.
Знаковое значение эмоций закладывается у низкоразвитых и формируется у развитых животных после рождения, у менее развитых – до рождения. Образуется задолго до предметной деятельности. Поэтому не имеет к ней никакого отношения. Это значит, что закон причинности легко игнорируется. Субъективное время эмоции – длительность. Полностью базируется на гомеостазе и обстоятельствах.
Движения выразительной мимики, пантомимы и пластики происходят под управлением бледного шара. Афферентный синтез и сенсорные коррекции движений происходят в промежуточном мозге. Изначально движения выражения эмоций не нуждаются в коррекции дистантной рецепцией. Но, по мере развития каналов информации между животными, коррекции от дистантной рецепторики возникают и развиваются. Выражение собственных эмоций происходит в присутствии другого, и этот другой, будучи особью того же вида будет реагировать на проявление эмоций выражением собственных эмоций. Через органы чувств вы получите информацию о состоянии Вашего визави, вы легко сделаете вывод об этом его состоянии и его соответствии Вашему, о вашей с собеседником способности и склонности к совместной деятельности.
Эти средства коммуникации в филогенезе становятся все более совершенными, Поэтому мы легко определяем эмоциональное состояние у млекопитающих, овладевших уровнем пространственного поля, и труднее – у более древних групп животных. Формирование эмоций происходит с учетом состояния внутренней среды организма, что подразумевает участие промежуточного мозга, управление выразительной мимикой согласно Бернштейну бледный шар (подкорковые ядра), за длительность эмоционального состояния ответственна первичная кора лимбической системы. Кроме состояния организма на эмоциональное состояние влияет внешняя обстановка – наличие или отсутствие опасности, пищи, особей своего вида своего и противоположного пола. (рис.7 гл 2)
На уровне эмоций, управляемых лимбической системой, течение времени иное, чем при действиях, управляемых иными структурами мозга. Если на уровне В время представляет собой ритм смены собственных состояний, на уровне С время длится от какого то начала до конца, на уровне Д закон причинности задается восприятием времени как «до» и «после», то на уровне эмоций время воспринимается единым куском от начала передачи информации от одной особи к другой до полного завершения всех эмоциональных реакций этого диалога. В это время оба собеседника как бы выпадают из прочих видов течения времени, они находятся в едином времени, временной точке, которая имеет размеры от начала до конца диалога.
Восприятие времени в это время совсем особое. Главное в отличии от иных способов восприятия времени в том, что, поскольку течение эмоциональных реакций в числе прочего имеет своей целью синхронизацию работы двух организмов, отслеживаются только те эмоциональные действия визави, которые требуют коррекции со стороны собственного организма. Не имеет значения на всем временном промежутке диалога, что было до и после какой-то локальной коррекции, значение имеет только полное время контакта, которое воспринимается одновременно и как длительность от начала до конца диалога, и как точка, мгновение, после которого судьба одного или обоих участников может резко изменить свое до сих пор ровное течение. В момент эмоционального контакта закон причинности игнорируется. Ведущее значение имеет состояние собственного тела и партнера.
В процессе воспитания животные осваивают выразительную мимику тех, кто рядом. Животные маугли так же известны науке, как и люди-Маугли. Овладев в раннем возрасте способами эмоционального общения с животными других видов, животное утрачивает способности нормального общения с животными своего вида. Это значит, что участки коры, ответственные за освоение языка эмоций после освоения одного языка не могут освоить другой, или, по крайней мере, в значительной степени утрачивают эту способность с возрастом. Мимические мышцы человека и млекопитающих управляются корой нижней части прецентральной извилины. Стало быть, эта зона коры находится под непосредственным влиянием лимбической системы. Мимика сосредоточения говорит об отсутствии этой связи в момент предметной деятельности. Т.е. предметная деятельность тормозит проявление эмоций в мимике, что в свою очередь означает, что уже на уровне предметных действий Д деятельность лобной коры в состоянии подавить, по крайне мере на время, действие лимбической системы. В это время (в момент сосредоточения на предметной деятельности) время осознается как до и после, длительности нет. Каждая элементарная предметная операция занимает строго определенное место во временной последовательности. При включении мимики, т.е. при выражении эмоций, течение времени для субъекта меняется, нет «до» и «после», есть длительность.
Эмоциональные состояния не могут меняться произвольно. Если дать эмоциям возможность свободно протекать, их последовательность быстро подстроится под ритмы внешнего мира. Точнее приведет ритмы организма в соответствие с ритмами мира. Сложность в том, что смена ритмов в самом организме протекает с разной скоростью. Действие опорно-двигательного аппарата опережает изменения в работе внутренних органов. Любое мышечное действие влечет за собой последовательность изменения работы внутренних органов, которая полностью заканчивается, иногда, через несколько суток после окончания работы мышц. Начатая последовательность эмоций должна отыграться полностью.
Организм не может не сказать Б если сказал А.
Механизмы выражения эмоций и чтения их у человека не слабее, чем у животных. Материальный субстрат присутствует в полном объеме. Мало того, механизм формирования языка эмоционального общения включается в онтогенезе автоматически, и у каждого он есть.
Человеческая речь – не вторая сигнальная система в филогенезе, а третья по счету сигнальная система. В филогенезе речь формируется после того, как произошли два параллельных длительно становящихся процесса – становление эмоциональной сигнальной системы (при помощи развития в ЦНС лимбической системы), и становление предметной деятельности (развитие третичной коры и в особенности лобных долей головного мозга). Оба процесса доведены до человеческого уровня, освоены и сформированы и генетически закреплены в коре центры, ответственные за каждую из них. Следовательно, на уровне эмоций течение времени в каждом контакте становится одной большой временной точкой, а на предметном уровне сформировано точное деление времени на «до и после» каждого конкретного действия, имеющего точно определенные начало и конец. Каждый звук в слове и каждое слово в предложении занимают свое строго определенное место.
Таким образом, мы имеем три сигнальных системы. Если расположить их в порядке возникновения в филогенезе, это первая сигнальная система – анализаторы, формирующие понятия собственного тела, пространственного поля, предмета и, апофеоз этого уровня – закон причинности; эмоциональная сигнальная система, возникшая едва ли не раньше первой, и базирующаяся на лимбической системе и вторая сигнальная система, полностью базирующаяся на третичной молодой коре головного мозга. Первая сигнальная система ориентирует организм в окружающей обстановке, эмоциональная сигнальная система ориентирует организм в его жизненных потребностях и взаимоотношениях с особями своего вида, вторая сигнальная система целиком построенная на взаимоотношениях с особями своего вида есть использование первой сигнальной системой в интересах эмоциональной сигнальной системы.
Поэтому вторая сигнальная система легче повреждается и труднее восстанавливается, чем эмоциональная сигнальная система. Эмоциональная сигнальная система имеет более древнюю базу, и наследственные механизмы формирования. Дети научаются понимать эмоции родителей и реагировать на них в соответствии с правилами общения людей раньше, чем научаются говорить.
Мы при общении используем обе системы — эмоциональную и речь. Речь в чистом виде – универсальнее, т.к. есть слова – символы эмоций. Они действуют сразу на две системы. Эмоции можно скрыть за словами. Тогда общение становится опасным для искреннего человека.
2.6 СОЧЕТАННОЕ РАЗВИТИЕ ПРЕДМЕТНОЙ ДЕЯТЕЛЬНОСТИ И ЭМОЦИОНАЛЬНОЙ СИГНАЛЬНОЙ СИСТЕМЫ.
Усложнение мозга в процессе антропогенеза шло по двум параллельным линиям:
• от управления локомоциями, через предметную деятельность и символические действия к речи;
• от формирования лимбической доминанты в стае, через развитие эмоциональной системы общения, через развитие мимико-звуковой системы общения к речи.
Развитие коры головного мозга берет начало от обонятельного мозга. Древнейшая кора возникла и всегда связана с обонятельным рецептором. Возникновение этого отдела мозга на переднем конце тела и обусловлено расположением здесь хеморецепторов. У первых хордовых движения на этом уровне полностью управляются спинным мозгом. Чувствительная кора обонятельного анализатора фактически ориентирует организм в пространстве относительно градиента концентрации привлекательного химического вещества. Таким образом, основными функциями древнейшей коры было управление намерением живого существа – выбор направления движения, устойчивость желания, учет гомеостаза. Первичная кора делала афферентный синтез, создавая картину мира, в состав которой входил привлекательный образ внешней среды, и состояние собственного организма. Огромное значение это имело в половом размножении — хемотаксис по направлению к самке. Первичная кора имела функцию чувствительного анализатора, роль двигательной коры она не выполняла. Процессы, происходящие в первичной коре, были относительно самостоятельны – прямо не влияли ни на движения тела, но по мере влияния внешних и внутренних условий влияли на стремление тела к пище и особи противоположного пола.
В то же время, сложность строения коры, избыточное количество нейронов в ней по сравнению с древнейшими образованиями создали условия для движения возбуждения по замкнутому кольцу, в течение длительного времени, обусловили эффект последействия и суммирования эффектов во времени,что делало процессы в первичной коре относительно автономными по отношению к процессам, происходящим в организме. Циклические процессы в первичной коре в силу своей электромагнитной природы создавали одноименные волны, которые распространялись по всем направлениям со скоростью света, и сила их изменялась обратно пропорционально квадрату расстояния от источника.
Большая часть, если не вся первичная кора была представлена вставочными нейронами, большинство из которых было соединено со вставочными же нейронами. Синаптическая система передачи нервных импульсов была несовершенна, и нейроны первичной коры реагировали не только на сигнал, полученный через синапс, но и на изменения электрического поля. Чувствительность нейронов к переменным электромагнитным полям определяла чувствительность коры к источникам этих полей. Помимо не биологических источников, в природе достаточно много биологических источников. Одним из главных свойств коры, а тем более, первичной, было усвоение ритма. Источники наиболее близких ритмов – особи того же вида. Особи одного вида чувствовали друг друга, как при помощи хеморецепции, так и при помощи полевой чувствительности. Полевая чувствительность дублировалась хеморецепцией, и оба вида чувствительности дополняли друг друга, и замыкались на первичной коре. У человека древнейшая кора обонятельного мозга представлена лимбической системой. Для полового поведения и продолжения рода и для поиска пищи это надежнейшая связь.
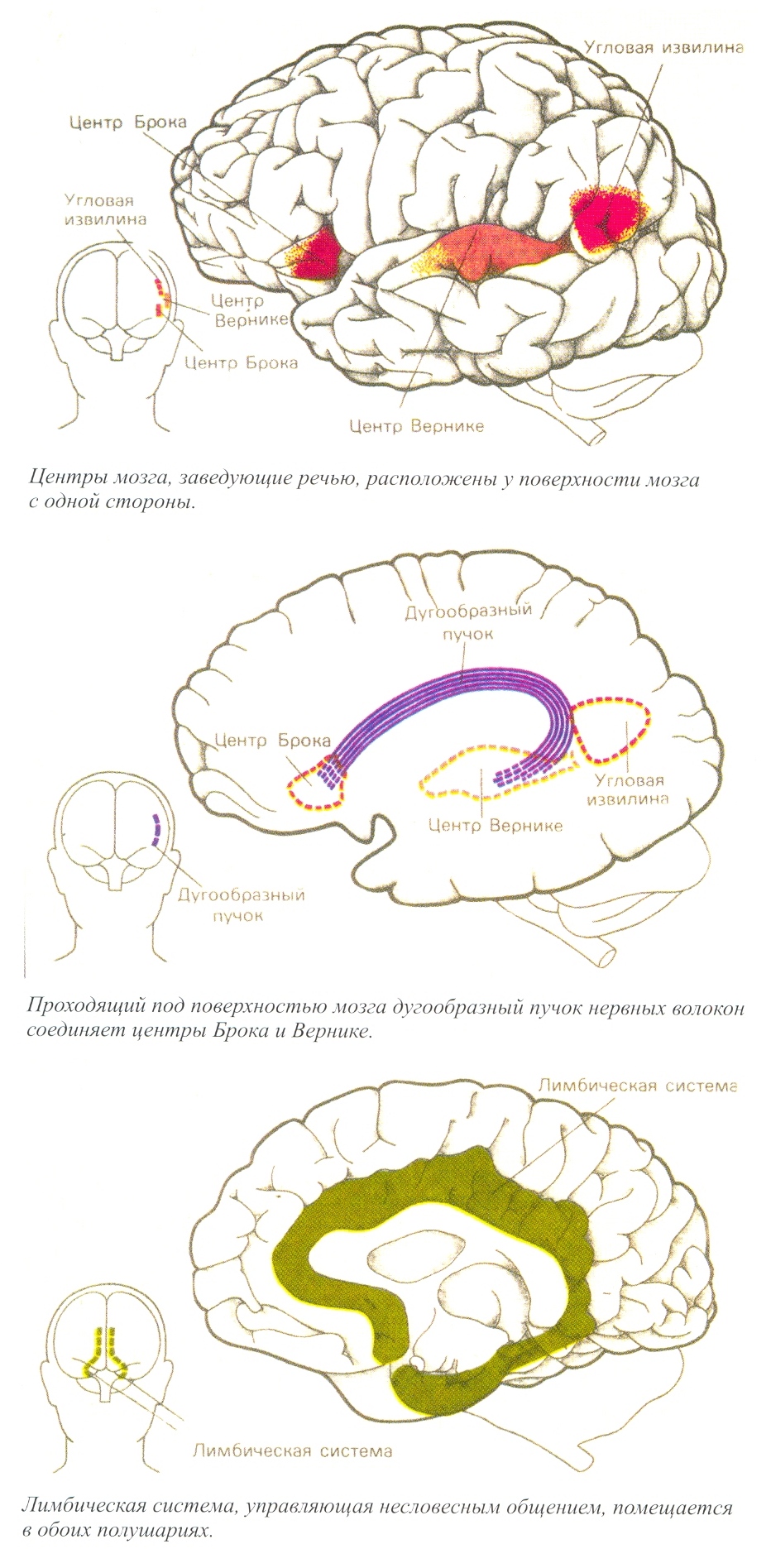
Если хеморецепция обеспечивала направление движения к особи противоположного пола, то наличие полевой чувствительности позволяло синхронизировать особи одного вида. Такая синхронизация обеспечила возникновение стаи. Для стаи характерно совместное действие всех особей одного вида. Каждый мог наблюдать такое действие, наблюдая стаю рыб или птиц, поражаясь синхронности их действий. Действительно, стая действует как единый организм, и она и есть единый организм в тот момент, так как работа первичной коры каждой особи синхронизирована с первичной корой каждого соседа. Стая, действуя как единый организм, снижает вероятность удачной атаки хищников, резко повышает выживаемость вида. Этот факт и обеспечил продолжение жизни у получивших обонятельный мозг – первичную кору. Здесь необходимо заметить, что при дальнейшем усложнении нервной системы у позвоночных, другие дистантные органы чувств – зрение, слух, органы боковой линии у рыб изначально не имели представительства в коре. Зрение и слух имели представительство в среднем мозге. Это было обусловлено тем, что для первых позвоночных, небольших по размеру, при обилии уже хорошо развитых хищников другой эволюционной направленности, очень важно было спастись от гибели. Зрение и слух, в отличие от обоняния, не вели к цели. Эти органы сигнализировали об опасности, предупреждали заранее, о том, что опасность близка, необходимо в настоящий момент, прямо сейчас, все бросить, и бежать, подальше от этого места. Именно средний мозг вполне успешно конкурирует с корой головного мозга за конечные двигательные пути при опасности даже и у человека. Часть нервной трубки, соответствующая у первых хордовых среднему мозгу, будучи расположена ближе к двигательным нейронам, чем обонятельный мозг и была ответственна за принятие решения о выборе управления, а следовательно, и направления – то ли стремиться к еде и самке, то ли бежать от опасности.
Постепенно все большее количество органов чувств получало представительство в коре. Под стать органам была и кора. Зрительная и слуховая кора были заняты, прежде всего, оценкой степени опасности, предвещаемой сигналами. Подкорковые центры зрения и слуха были в этом достаточно совершенны, кора, получающая сигналы от этих органов чувств была освобождена от этой утилитарной работы. Ее деятельность носила все более сигнальный характер. Каждый сигнал, полученный корой головного мозга по этим каналам, уже на уровне среднего мозга освобождался от оценки о непосредственной опасности. Теперь можно было оценить информацию не особенно спеша, на предмет дальних предсказаний. Причем сигналы, полученные корой от дистантных рецепторов можно было сопоставить с сигналами обонятельной и вкусовой коры, с информацией от опорно-двигательного аппарата, от внутренней среды организма. Иначе говоря, по мере развития коры и двигательных возможностей, формирование картины внешнего мира становилось все более полным. Бернштейн называет афферентный синтез этого уровня пространственным полем. Лимбическая система в это поле помещает ту самую цель, к которой совершает локомоции позвоночный организм. Из чисто обонятельной и вкусовой коры появляется центр личности, которая не только боится умереть, но и хочет жить. Умереть боится на уровне среднего мозга, а жить хочет ведущей к цели корой головного мозга, той ее частью, которая формирует эмоции, а с ними и интерес к жизни. Позвоночный организм имеет цель, к которой у организма есть личный интерес, поддержанный циклическими процессами на уровне лимбической системы. Лимбическая система при этом еще и учитывает информацию, полученную по полевому каналу, то есть, прежде всего от особей своего вида. Таким образом, бывший обонятельный мозг становится центром личности, учитывает все, и все больше выходит из реального времени пространственного поля. Ведь учет состояния партнеров требует именно выхода из необходимости непосредственно реагировать на всю остальную обстановку в пространственном поле – вот сейчас партнер выругается за то, что его отвлекли от дела, одумается и поймет, что ему только что передали жизненно важную информацию. Реакция партнера будет отсрочена во времени, но для обоих партнеров этот эмоционально насыщенный промежуток времени воспринимается как мгновение, хотя длиться реально может несколько минут. Отсюда возникает требование анатомического разделения органов, управляющих течением времени контакта между личностями, и органов, принимающих на себя управление движениями, когда это время истекло и пора выполнять уже принятое решение. Таким образом, функционально, а затем и анатомически выделилась лобная доля, с ее пирамидной системой, а затем и зоной Пенфилда.
Итак, появилась лобная доля. В коре полушарий появились функционально определенные зоны — так произошла функциональная дифференциация коры а вместе с ней и возможность непосредственно управлять деятельностью организма в окружающей среде.
Стайное поведение сохраняется на всем протяжении филогенеза позвоночных – от рыб до приматов. По мере совершенствования лимбической системы совершенствуется и групповое поведение. Возникновение рефлекса подражания имеет происхождение из полевой чувствительности предшественников лимбической системы – древней обонятельной коры. Дело в том, что стайное поведение рыб, когда, реагируя на изменение обстановки, вся стая, как одно существо, состоящее из отдельных рыбок, как организм состоит из клеток, совершает синхронное действие, все это обусловлено тем, что каждая рыбка чувствует поле всех остальных и изменения в суммарном, групповом поле не являются секретом для каждой рыбки. Мысли одной особи неотличимы от мыслей любой другой. Общее поле, создаваемое специализированными клетками, способными накапливать заряд и сохранять на мембране разность потенциалов, является для тех же клеток сигнальным, сообщением об изменении обстановки в группе. Это поле связывает особей одного вида в коллектив. В этом коллективе каждая особь для любой другой есть частица, принимающая участие в создании общего поля. Друг для друга все особи безлики. При стремлении к пище и спасении от опасности не имеет значения, кто конкретно рядом с тобой. Но все меняется, когда наступает время спаривания. Здесь не должно быть ошибки – партнер должен быть противоположного пола, должен быть половозрелым и степень его готовности к спариванию соответствовала вашей. Некоторая разница в непосредственной готовности устранялась при так называемых брачных играх – эти игры имели в то время и для тех видов именно этот смысл. В силу того, что зрение, слух, проприорецепция при этом имели малое значение, главную роль при опознании подходящего партнера играла система, у млекопитающих развившаяся в лимбическую. При половом поведении первоочередное значение имеет собственное состояние, состояние гомеостаза, опознание партнера, и аналогичное состояние у партнера. Все это делало совместное действие эффективным, способствовало продолжению рода, и, будучи у высших позвоночных усовершенствовано выбором наилучшего из партнеров, было закреплено в терминах хромосом.
Одним из примеров того, что хеморецепция и обонятельный мозг тесно связывает пищевое и половое поведение, дает пример с поведением акул, при пожирании стаей большой легкодоступной добычи – мертвого кита. Акулы настолько увлекаются процессом группового питания, что он у акул легко переходит в процесс совокупления с особями противоположного пола.
Но речь здесь немного о другом – о тесной связи между сотрудничеством и лимбической системой. Именно этот отдел головного мозга ответствен за активное поведение в окружающей среде, за пищевое и половое поведение, за прием и передачу электромагнитных сигналов в окружающую среду и за формирование эмоций. На уровне рыб химические и электромагнитные сигналы остаются практически главными средствами коммуникации, у змей мы можем наблюдать брачные танцы, у лягушек – звуки, у птиц – сложное пение и танцы. Но современных видов рыб и у птиц мы наблюдаем сложные формы парного сотрудничества в деле выращивания потомства. По крайней мере, в птичьих парах мы видим, что это сотрудничество подразумевает достаточно длительные взаимоотношения между особями одного вида, основанные на выращивании потомства. Видимые нами средства коммуникации не позволяют в достаточной степени договориться. Многое мы можем списать на врожденные программы действий, в том числе и совместных. Но процедура такого списания ставит перед нами вопрос о том, откуда, как, зачем и почему у другой линии теплокровных – у млекопитающих развились мимические мышцы, и все в их поведении, связанное с выражением эмоций.
Древняя кора обеспечивает устойчивость эмоций. Её тесная связь с промежуточным мозгом ставит операции, проводимые с её участием, зависимыми от гомеостаза. Её влияние на поведенческие реакции (взаимоотношения с другими особями) указывает на ее связь с мимическими действиями. Древняя кора задаёт тон при подражании – именно этот эффект связан с обучением. Невозможно объяснить появление и развитие мимической мускулатуры у млекопитающих без учета явления подражания. Но само подражание основано на особом канале коммуникации – электромагнитном поле, и в первую очередь производимое древней корой.
Появление мимической сигнальной системы означает выделение из всей мускулатуры особой части, ответственной только за общение с себе подобными.
При помощи мимики передается другому эмоциональное состояние, что в свою очередь есть интегральное выражение гомеостаза, физиологического состояния организма, оценки окружающей обстановки, и отношение к этому другому – его рассматривают, исходя из вышеперечисленного, как полового партнера, как партнера по охоте, как партнера по самозащите, по игре, как соперника, как врага, друга, брата и т.д.
Каждое из рассмотренных эмоциональных отношений предполагает соответствующее ответное действие. Да, я согласен, нет мимической системы ни у рыб, птиц, земноводных, тем не менее, они так же могут совершать совместные действия. Но это и является подтверждением того, что и они имеют структуры, ответственные за совместные действия. Лимбическая система и ее аналоги связывают отдельные особи в стаи, пары. Она же позволяет юному потомству подражать действиям родителей. Получая от собеседника полевой сигнал, невозможно отличить его от сигналов собственного тела. Собственное тело само продуцирует подобные сигналы, и о чем собственно это сообщение. Это сообщение, передаваемое электромагнитные сигналы от древней коры, говорит о состоянии организма. Оно говорит об эмоциональном отношении к состоянию собственного тела и окружающей его обстановке. Получив такое сообщение, вы можете его совсем не заметить, если Ваше личное состояние полностью соответствует полученной информации. Оценка обстановки может быть различной, но если Вашего партнера что-то в ней беспокоит, то вы, восприняв это беспокойство можете забеспокоиться по тому же поводу, но можете понять как причину своего беспокойства возможную опасность или наоборот, возможность появления в обстановке чего то привлекательного, и у Вас не будет повода считать это беспокойство как полученный сигнал. Вы этот сигнал примете как изменение собственного состояния. Если же в вашем теле есть больное место, то оно будет вас беспокоить, вносить свой вклад в ваше эмоциональное состояние, и ваша древняя кора передаст этот сигнал всем присутствующим в пределах досягаемости поля. В этом случае сигнал о чьем-то добром здоровье принятый вами будет сильно отличаться от вашего эмоционального состояния, вы легко почувствуете разницу – только что было очень плохо, и вот стало немного лучше. Таким образом, в стае каждая особь чувствует себя как средняя особь в стае. Выпасть из стаи – значит погибнуть – ведь вокруг есть и хищники, которые вполне могут использовать индуцируемую вами информацию о вашем состоянии.
Последствия полового акта для особей противоположного пола различны. Особи мужского пола не могут чувствовать в себе изменений, происходящих в женском организме. Полевая информация говорит мужчинам о чем то необычном, не более. Особи женского пола имеют готовую биологическую программу действий в случае наступления беременности. Договориться о дальнейшем сотрудничестве в этом случае гораздо сложнее, чем совершить половой акт. Если у низших позвоночных в этом случае полевая работа древней коры дополняется врожденными поведенческими реакциями, то у млекопитающих, вследствие развития старой коры сигналы древней становятся все более приглушенными. По мере эволюционного роста млекопитающих, древняя кора имеет все меньший удельный вес в общей массе коры головного мозга, и сигналы древней коры все более заглушаются сигналами молодой коры. Эта же молодая кора связана опять же в эволюционном росте все более с окружающим пространством, с афферентным синтезом пространственного поля, а не собственного тела, а заодно и духа. В природе эта проблема решается параллельно двумя способами. Во-первых, на базе древней коры формируется лимбическая система, представляющая два параллельных кольца, соединенных между собой особыми образованиями (свод, мозолистое тело, передняя спайка), внутри которых расположены как стержень части боковых желудочков, заполненных ликвором. Эта система не только позволяет длительную самоподдерживающуюся циркуляцию импульсов, что обеспечивает устойчивость эмоциональных состояний, но будучи взяты как единая конструкция, представляют собой антенну для приема и передачи электромагнитных волн. Таким образом, одна и та же система и формирует эмоциональное отношение к себе и к миру, и является средством коммуникации между особями. Причем связь между биологическими особями происходит одновременно по двум каналам – полевой канал, прямая связь между лимбическими системами двух особей, и канал демонстрации эмоций через движения тела.
Здесь то и таится возможность развития речи в будущем. Дело в том, что если бы канал связи был один, то все усовершенствования этого канала закреплялись бы в хромосомном аппарате, и пользование таким каналом не требовало бы научения в постнатальном онтогенезе. А возникновение второго канала – проявление состояния тела в движениях, а впоследствии и в выразительной мимике было бы излишним. Но цефализация привела к тому, что жизненно важный синхронизатор совместной деятельности двух особей – древняя кора – становился все более закрыт. Появление и развитие, разрастание новой коры привело к тому, что лимбическая антенна экранировалась новой корой. Новая же кора по своей функции формировала сигналы нестабильные, как по месту их возникновения, так и по ритмам. Сигналы получались огромной сложности, и, в силу этого, невнятными. Воспринять их как принимающая антенна кора головного мозга может, но отделить свои собственные токи от токов, наведенных другой корой, принятых из электромагнитного поля, для новой коры сложно. Исходя из этого, и возникла необходимость в ином канале связи. Этот канал проявил себя в том, что состояния организма выражаются в движениях. Больной организм движется иначе, чем здоровый, эмоциональное состояние проявляется достаточно явно в рисунке движений. Уже при появлении конечностей достаточной длины появилась возможность собственными конечностями воздействовать на части собственного организма. Для движений обслуживания своего тела, птицам достаточно развитых базальных ядер. Но сама необходимость таких движений, их частота, интенсивность, размах и уместность могут быть для наблюдателя достаточно информативны. Наиболее же важными и информативными являются движения переднего конца тела, и особенно тех его частей, какие ответственны за наиболее жизненные функции – пищевое поведение, дыхание, и ориентирование в пространстве. Рот, глаза, уши, по меньшей мере их направление, степень сосредоточения многое говорят опытному глазу об эмоциональном состоянии. Положение конечностей, тела, поза в целом, движения хвоста, возможно отвлекающие, могут сказать о намерениях. Тонус мышц, энергия в движениях и в позе говорят о состоянии здоровья. А собственная развитая кора, способная к анализу, легко прочитает все эти знаки. Если имеет такой опыт. Я имею в виду опыт чтения знаков тела. Конечно он может быть приобретен предками, и записан в хромосомах, но тогда не надо обижаться на себя и предков за неправильный учет всей обстановки в целом.
Древняя кора раньше новой созревает в онтогенезе, и пока новая кора еще не сформирована, ее сигналы не очень мешают настроиться на сигналы лимбической системы матери. Умение читать эмоциональное состояние мамы и должно быть, и оно есть у новорожденных, скорее всего как врожденное. Канал лимбической полевой связи между мамой и младенцем начинает работать сразу. По мере развития новой коры, в ней устанавливаются связи между сигналами, полученными по полевому каналу, и по каналам первой сигнальной системы (по Павлову). Таким образом врожденные реакции лимбической системы младенца на сигналы такой же системы матери сочетаются с сигналами, получаемыми от дистантных органов чувств. Так происходит обучение новой коры восприятию эмоций матери по ее внешне выраженным эмоциональным реакциям. Итогом такого обучения становится совершенно аналогичное выражение эмоций младенцем.
И какая же мать согласится отдать своего дорогого ребенка – лягушонка, слоненка, тигренка в обучение кому – то другому. В этом случае, выражение эмоций ребенком будет зависеть от учителя. Если же ребенок не способен воспринимать по каналам первой сигнальной системы, он воспримет эмоции матери от нее же по полевому каналу, и по этому каналу и будет происходить обучение. Даже слепые от рождения дети, никогда не видевшие мамы, точно воспроизводят ее мимику. Все равно, их древняя кора обучает новую, но только по своим каналам, используя не зрение, а электромагнитные сигналы лимбической системы, вводя окружающих в соблазн мракобесия.
Таким образом, у млекопитающих по каналам первой сигнальной системы передается информация от одной особи к другой об эмоциональном состоянии. Причем для передачи информации используются движения тела, производящие жесты, мимику, звуки, а для приема информации используется зрение и слух. Дублирующим каналом является полевая связь между лимбическими системами двух особей. В случае полевой связи лимбическая система работает и на прием и на передачу.
Оба эти канала передачи и приема эмоционального состояния от одной особи к другой мы объединим одним названием – эмоциональная сигнальная система. Полевая связь здесь древнейшая. Затем в порядке появления идут позы, жесты, затем звуки, затем мимика. Жесты, то есть движения телом, читаются вначале при помощи среднего мозга, промежуточного, а затем и при помощи коры зрительного анализатора. Именно поэтому зрительные бугры и получили преимущество, став местом синтеза всей телесной информации. Зрительный же анализатор, задолго до того, как стал участником афферентного синтеза пространственного поля и предмета, давно уже был участником эмоциональной сигнальной системы. Вначале слабым, вспомогательным, а затем все более сильным и полноправным. Вначале как дополнительный сигнал, уточняющий информацию, полученную по полевому каналу, а затем как вполне самостоятельный. То же можно сказать и о слуховом анализаторе. Афферентный синтез эмоциональной сигнальной системы – партнерство, отношения.
Таким образом, через движения, т.е. управлением сокращения мышц опорно-двигательного аппарата происходит выражение эмоционального состояния. Управления движениями согласно Бернштейну происходят у человека с уровней А, В, С, Д и Е. На всех уровнях лимбическая система вносит свой вклад в управление движениями. На каждом уровне вклад лимбической системы различен у разных видов. Это связано с тем, что собственный вклад каждого уровня в управление движениями у разных видов различен. У рыб обонятельный мозг задает направление движениям, тропизм и таксис к пище и особям противоположного пола, а так же и управляет движениями, связанными с групповым поведением, а поскольку рыбий потолок – В, то на этом уровне все и происходит. У человека уровень В несет ответственность за синергии мышц. Для нас этот уровень интересен тем, что на нём происходит управление движениями эмоционального общения – объятия, поцелуи, ласки. Таким образом, кожная и проприоцептивная чувствительность делают свой вклад в эмоциональное общение двух личностей. То есть движения, управление которыми происходит от бледного шара, дают отклик в организме: во первых – проприоцептивная обратная афферентация от собственных органов движения, во вторых – кожная чувствительность от соприкосновений с телом партнера, которая разделяется на ощущения от соприкосновений от собственных действий, и на ощущения от соприкосновений от действий партнера. К этому добавляются ощущения от реакций внутренних органов на объятия, поцелуи и ласки – те еще ощущения, это в третьих. И четвертое – полевые сигналы от лимбической системы. Афферентный синтез от рецепторов происходит на уровне зрительных бугров, создает картину событий в собственном теле, полевые ощущения воспринимаются на уровне древней коры, и там же формируется афферентный синтез партнера. Потому-то запахи имеют такое значение для опознания партнера и у животных и у человека. Древняя кора же принимает готовую картину собственного тела от промежуточного мозга, и образ партнера неотделим от образа собственного тела на этом уровне. Образ партнера дополняется показателями собственного гомеостаза. Потеря постоянного многолетнего партнера для животного означает на уровне лимбической системы потерю части самого себя, создает субъективное ощущение нехватки чего-то важного, без чего абсолютно все телесные ощущения и картина собственного тела становится неполной.
Иное происходит при управлении движениями на уровне С. Н.А. Бернштейн разделяет этот уровень на два подуровня – С1 и С2. Афферентный синтез уровня С – пространственное поле. Движения на этом уровне имеют выраженный целевой характер. Жизненно важные цели определяет опять же лимбическая система. Объекты пищевого и полового интереса это живые существа. Афферентный синтез системы распознавания живое – неживое, привлекательно -непривлекательно, не может обойтись без лимбической системы. Распознавание опасно-неопасно происходит без её участия (на уровне среднего мозга). В состав движений уровня С Бернштейн включает и не только чисто локомоторные, переместительные и целевые движения. Сюда же он относит и такие движения, как подражательные, копирующие, мимические, выразительные движения, педагогические движения, движения заботы, в том числе и кормление. Но все эти движения есть движения не сами по себе, и, даже если они несут в себе локомоторный компонент, то они при этом обязательно подразумевают наличие как цели другого живого существа, как правило, своего вида, или детеныша, или подразумевают наблюдателя, которому посылается сигнал, информация, демонстрация собственной личности для межличностного взаимодействия.
Подражать кому-то мы можем, если этот кто-то уже воспринимается нами как нам подобный, чье состояние в данный момент нам интересно, кого мы хотим заинтересовать собой. Если мы хотим перенять его опыт в движениях, подражая ему, мы проводим сопоставление по двум каналам не только частям его тела свои части тела, но и состояние своей лимбической системы с таковым у объекта подражания. Иначе не будет никакого обучения, мы не сможем определить, правильно мы делаем движение или нет. Наш партнер – образец — нам подскажет. Педагогические движения, само собой разумеется, имеют отношение к потомству. И здесь тоже невозможно представить себе действенность показа действий без параллельного воздействия на обучаемого через полевой канал. Движения обслуживания тела детеныша и его кормление тесно связаны между собой, и прежде всего, опять же связь детей и родителей уже существует, уже действует тот невидимый канал. Но наибольшее проявление находит взаимодействие двух личностей при появлении символических движений с помощью мимической мускулатуры.
Мимическая мускулатура есть производное от мышц лицевой части головы. Это мышцы, обеспечивающие открывание и закрывание глаз, рта, носа. Собственно мимические мышцы появляются только у млекопитающих, эти мышцы изменяют конфигурацию мягких тканей головы и лица. Представьте себе изменение выражения лица у лягушки, голубя, крокодила или змеи. Что вы можете прочесть на их лицах? Ничего, и это следствие явного недоразвития, точнее отсутствия мимических мышц у этих видов. Но и млекопитающие наделены этой мускулатурой в разной степени. Наиболее сильно мимические мышцы развиты у приматов, и самое сильное развитие получили у человека. То есть, человек в процессе антропогенеза помимо прочего тренировал еще и мимические мышцы. И не просто так тренировал, а тот, у кого они были сильнее развиты, получил преимущество при размножении. Это не гипотеза, это факт. Само по себе прогрессивное развитие мимической мускулатуры происходило параллельно с антропогенезом. Антропогенез сопровождался усилением коммуникаций при посредстве символических действий, производимых мышцами лица. Чем амимичное лицо хуже лица с выразительной мимикой? Тем, что при коммуникациях партнеру приходится опираться либо на полевой канал, либо на жесты и позы. Но полевой канал мешает отделить собственное состояние от состояния партнера, полевой канал может обеспечить только стайное поведение. Для более тонкого взаимодействия, когда каждый из партнеров четко отделяет себя от другого, что позволяет легко разделить функции при совместном действии, полевой канал не подходит. Канал выразительных поз и жестов позволяет согласовать многие действия у достаточно продвинутых млекопитающих, при этом парные действия, связанные с функцией размножения, в значительной степени обеспечены врожденными программами соответствующих действий. У приматов, особенно прямоходящих, имеющих достаточно сильные ограничения в размножении, установление межличностного контакта при размножении становится первостепенной необходимостью. Недостаточно вступить в половой контакт, недостаточно при этом выбрать биологически полноценного партнера, надо, что бы этот партнер еще и был достаточно ответственным при беременности, родах и вскармливании младенца, которое длится 1-2-3 года, обеспечивал бы пропитание матери, в период её беспомощности, обеспечение всем необходимым для жизни, а список необходимого все растет . Кроме того, как мы уже увидели выше, неандертальцы произошли от австралопитеков, живших в домах, носивших одежду, пользовавшихся огнем, овладевающих уровнем Д (предметных действий). Умения неандертальцев уже были достаточно обширны, для того, чтобы обучение этим умениям было серьёзной работой и для учителей, и для учеников. Все это требует длительных коммуникативных взаимодействий не только при размножении, но и после того, как дети родились, выросли, и у них возникла необходимость построить свой дом, а речи то еще нет. Отсюда максимальное использование и постоянное развитие имеющихся каналов коммуникации. А этих каналов два – полевой с использованием лимбической системы, и канал с использованием движений тела. Мимическая мускулатура стала отличным дополнением к позам и жестам, она же стала необходимым шагом в развитии действий символических, совершенствуя уровень Д до предела, допустимого для неандертальцев. Управляя мышцами лица, неандертальцы научились передавать достаточно тонкие оттенки эмоций. Тем самым убивались два зайца: действия минимизировались энергетически, и общение стало очень личностным.
Минимизация энергозатрат при общении привела к возможности избыточных действий. Совсем не трудно еще и еще раз повторить действие, не требующее использования силы. Возможно общение при положении тела в спокойном состоянии. Можно, обратив лицо к собеседнику, не прерывая работы, продемонстрировать ему свое состояние как эмоциональное, так и состояние здоровья, можно показать свою оценку съедаемого или только что съеденного продукта, среагировать на события в семье, показав отношение к ним, но не вмешиваясь в их ход действием. Постоянная, по любому поводу игра мимическими мышцами (а это был помимо прочего еще и уровень предметных действий, символическое действие) развивала уровень Д – именно его символическую сторону, тренируя мозг в направлении знаковых действий, малоэнергетичных, эмоционально заряженных, выражающих отношение каждого к себе, миру, а, главное – к другим личностям. Неандертальцы не могли говорить словами, они говорили позами, жестами, мимикой и лимбической системой. Они относились друг к другу как собеседники, то есть выделяли тех, кто их понимал, от тех, кто их не понимал, или понимал, но недостаточно. Говорить они не могли.
Усложнение мозга шло по двум параллельным линиям:
-
от локомоций, через предметную деятельность и операции с символами к речи;
-
от формирования лимбической доминанты в стае, через развитие эмоциональной системы общения, через развитие мимико-звуковой системы общения к речи.
На протяжении антропогенеза шла конкуренция между памятью хромосомной и памятью ЦНС. Хромосомная память при похолоданиях подавлялась, преимущество переходило к мозговой памяти, и наоборот, при спокойном течении жизни память переходила к хромосомам.
Центральная нервная система обладает специальными неспецифическими аппаратами, наделяющими доминантностью любую целостную деятельность, которая обеспечивает приспособительный успех организма в данный момент и про данных условиях. (Анохин). Концепция доминанты была предложена русским физиологом Ухтомским. Вот основные положения этой концепции, в виде цитат из Ухтомского.
Аппарат, представляющий собой множество переменных, калейдоскопически сменяющихся органов предупредительного восприятия, предвкушения и проектирования среды, и есть головной мозг. Процесс же смены действующих органов достигается посредством образования доминанты и торможения прочего мозгового поля.
Господствующий очаг возбуждения, предопределяющий в значительной степени характер текущих реакций центров в данный момент, обозначается термином доминанта.
Под именем доминанты понимается более или менее устойчивый очаг повышенной возбудимости центров, чем бы он ни был вызван, причем вновь привходящие в центры возбуждения служат усилению (подтверждению) возбуждения в очаге, тогда как в прочей центральной нервной системе широко разлиты явления торможения.
Нормальное отправление органа (например, нервного центра) в организме есть не предопределенное, раз навсегда неизменное качество данного органа, но функция от его состояния.
Главенствующее возбуждение организма в данный момент существенно изменяет роль некоторых центров и исходящих из них импульсов для данного момента.
Приписывание топографически определенному нервному центру всегда одной и той же неизменной функции есть лишь допущение, делаемое ради простоты рассуждения.
Внешним выражением доминанты является стационарно поддерживаемая работа или рабочая поза организма.
Концентрация внимания усиливается при возбуждении дыхательного и сосудистого центра.
3 фазы:
1). наметившаяся устойчивая доминанта привлекает в качестве поводов к возбуждению разнообразные рецепции (расширяется рецепторное поле);
2). из множества действующих рецепций доминанта вылавливает группу рецепций, которая для нее в особенности биологически интересна;
3). между доминантой и данным рецептивным содержанием устанавливается прочная связь, так что каждый из контрагентов будет вызывать и подкреплять исключительно друг друга, тогда как прочая душевная жизнь перейдет к новым текущим задачам и новообразованиям.
Среда поделилась целиком на предметы, каждому из которых отвечает определенная, однажды пережитая доминанта в организме, определенный биологический интерес прошлого. Я узнаю вновь внешние предметы, насколько воспроизвожу в себе прежние доминанты, и воспроизвожу мои доминанты, насколько узнаю соответствующие предметы среды.
Доминанта оставляет за собою в ЦНС прочный, иногда неизгладимый след. В душе могут жить одновременно множество потенциальных доминант – следов от прежней жизнедеятельности. Они поочередно выплывают в поле душевной работы и ясного внимания, живут здесь некоторое время, подводя свои итоги, и и затем снова погружаются вглубь, уступая поле товаркам. Но и при погружении из поля ясной работы сознания они не замирают и не прекращают своей жизни. Научные искания и намечающиеся мысли продолжают обогащаться, преобразовываться, расти и там, так будто возвратившись потом в сознание, они оказываются более содержательными, созревшими и обоснованными. Несколько сложных научных проблем могут зреть в подсознательном рядом и одновременно, лишь изредка выплывая в поле внимания, чтобы от времени до времени подвести свои итоги.
Заторможенные содержания пережитых доминант могут действовать патогенно, когда они не были в свое время достаточно вплетены и координированы в прочей психической массе. Тогда последующая душевная жизнь будет борьбою вытесняющих друг друга, несогласных доминант, которые стоят друг перед другом как инородные тела.
Доминанта – констелляция центров с повышенной возбудимостью в разнообразных этажах головного и спинного мозга, а так же в автономной системе.
Когда кора возобновляет прежде пережитую доминанту, дело идет о более или менее подробном восстановлении в организме всего комплекса центральных, мышечных, выделительных и сосудистых явлений. Когда нужно, кора умеет восстановить прежнюю констелляцию до такой полноты, что переживается вновь конкретное содержание тогдашнего опыта до галлюцинации. Обычно восстановление прежде пережитых доминант лишь частичное, экономическое, в виде символов. В связи с этим и комплекс органов, участвующих в переживании восстановленной доминанты, будет сокращенным – может ограничиться одним кортикальным уровнем.
Чисто кортикальная доминанта – есть позднейший продукт экономической выработки. Кора – орган возобновления и краткого переживания прежних доминант с меньшей инерцией и с целью их экономического сочетания.
Пережитая доминанта при затухании не аннулируется, а тормозится до поры до времени. Тому же принципу доминанты подчинена высшая нервная деятельность, это совершенно явствует из обыденного наблюдения, что в ответ на один и тот же сложный раздражитель (например научный доклад) оппоненты, прежде чем разберутся, разряжаются сначала каждый своим, что в нем накопилось, так что реплика определяется сплошь и рядом не столько тем, что выслушано, и не ближайшим содержанием выслушанного, а давними событиями.
Человек является настоящей жертвой своих доминант везде, где отдается предубеждению, предвзятости; и еще хуже, когда он сам этого не замечает. Чтобы не быть жертвой доминанты, надо быть ее командиром. По возможности полная подотчетность своих доминант и стратегическое умение управлять ими – вот практически что нужно. Предопределено давней историей, а сейчас совершается по ничтожному поводу – одна из трагических тем Ф.М. Достоевского.
Вот по этим «отрыжкам» скрывающихся двигателей поведения узнавать заранее зачатки своих мотивов, чтобы вовремя противопоставить им более важное, занять пути другою доминантою, — таков рецепт. Доминанта есть ли непременно корковое явление? Для меня несомненно, что она может закладываться еще в мозговом стволе, но коре приходится тотчас с нею считаться, поскольку кора для каждого мгновения есть орган сопоставления того, что требует сейчас внешняя среда, с тем, что делается во внутреннем хозяйстве тела.
Состояние возбуждения в доминанте надо пока характеризовать совокупностью следующих признаков:
- повышенная возбудимость: порог возбудимости в центре, становящемся доминантным, должен быть, по крайней мере, равен по величине раздражителю, доносящемуся до него в виде дальней волны возбуждения;
- стойкость возбуждения: чтобы начавшееся под влиянием дальней волны возбуждение в доминанте могло, в свою очередь, влиять на ход реакции, возбуждение это должно быть не мимолетным во времени;
- способность суммировать возбуждение при данной силе и частоте приходящих волн;
- инерция, при которой значение дальних волн сказывается преимущественно в подбадривании и ускорении установившейся доминантной реакции в направлении ее разрешения.
Таким образом, формирование поведения организма в среде, как единого целого, определяется тем, что в ЦНС формируется функциональный центр, берущий на себя управление всем организмом в данный момент. Остальные подобные центры, обеспечивающие поведение в иных обстоятельствах, в это время либо не работают, либо выполняют подчиненную функцию, а составляющие их нейроны в это время могут быть заторможены, или принимать участие в формировании доминирующего поведения.
Концепция Ухтомского достаточно плодотворна, при том, что и сегодня не решен вопрос о многих особенностях доминанты. Даже определение доминанты не имеет достаточно четкой формулировки. В то же время, исходя из концепции Бернштейна, мы можем многое конкретизировать в понятии доминанты и, с точки зрения доминанты рассмотреть уровни управления движениями. Мы рассчитываем, что подобный подход поможет нам прояснить некоторые этапы антропогенеза, до сих пор находящиеся в тени исследований антропологов, используя термин доминанта в некоторой степени .
На заре позвоночной жизни, когда кора головного мозга представлена еще только одним обонятельным мозгом, доминирующим центром был промежуточный мозг. Здесь происходила рецепция показателей внутренней среды организма, отсюда шли управляющие импульсы, заставляющие организм двигаться. В водной среде, для обеспечения организма кислородом необходимо движение. (Посмотрите на рыб. Они всегда находятся в движении, пусть медленном, и даже не вызывающем перемещения организма, но фильтрация жабр водой происходит непрерывно). Этот центр управления работает непрерывно, даже в случае, когда необходимо управление из другого места, дыхательные движения продолжаются. У высших позвоночных центры дыхания и кровообращения расположены в продолговатом мозге. Управление кровообращением нашему сознанию недоступно, это удел вегетативной нервной системы. Дыхательные мышцы могут работать под контролем сознания, но, если этот контроль снят, то они работают все равно, в любом состоянии сознания, под контролем дыхательного центра.
Позывы питаться и размножаться требуют определиться с целью, расположенной во внешней среде. Поиск цели происходил при помощи обонятельного мозга. При доминировании коры организм, ведомый намерением питаться или размножаться, поддерживаемым процессами в обонятельном мозге, устремлялся к пище или особи противоположного пола. При появлении опасности организм стремился избежать её, при этом управление организмом брали на себя нервные центры среднего мозга, информированные об окружающей обстановке дистантными рецепторами. Промежуточный мозг и в том и в другом случае производил учет состояния внутренней среды, и по состоянию внутренней среды вносились поправки в интенсивность движения. Таким образом, каждый отдел головного мозга брал ведущую роль на себя при определенных обстоятельствах. Личность могла проявить себя в полной мере, заявить «я хочу» и применить возможности организма, и центр управления – обонятельный мозг с его зачатками древней коры. При опасности желания личности отступали на второй план, на первый план выходило то, что мы теперь называем инстинкт самосохранения. Организм, игнорируя все остальные интересы личности, совершал поступки, ведущие к его сохранению, личность вместе с древней корой отступала на второй план. Что именно заставляло ее делать так, личность не могла определить. Неявные, неопределимые, не осознаваемые сигналы, доходящие до обонятельной коры от подкорковых образований, (а это повышение мышечного тонуса, изменение гормонального фона, состояния внутренней среды организма) позднее оформились в состояние коры, которое мы теперь называем страхом. Страх – с одной стороны, и влечение с другой стороны заставляли организм совершать движения. В каждом из этих случаев – состояние покоя, стремление к цели, избегание опасности.
Во всех трех случаях, организм вел себя по разному. Либо он стремился к лакомому куску, зная чего он хочет, либо он уносился сломя голову от опасности, плохо понимая, что именно она из себя представляет, либо лениво шевелил плавниками, восстанавливая константы гомеостаза и наслаждаясь покоем. В первом случае его слабая кора четко формулировала себе в терминах кортикальных процессов цель, являя собой полноценный акцептор результата действия (Анохин П.К. «Биология и нейрофизиология условного рефлекса» М.1968). Во втором случае никакой цели в коре головного мозга сформулировано не было, акцептор результата отсутствовал, было хаотическое возбуждение, обусловленное сигналами из среднего мозга – «Опасность!» во-первых, и сигналами о резком изменении показателей гомеостаза в связи с внезапно начавшейся физической работой, во-вторых. В третьем случае весь организм находился в состоянии восстановления. Наилучшим образом это состояние дополнялось заполненным желудочно-кишечным трактом.
Первый, из рассмотренных выше случаев, при наличии особей своего вида и противоположного пола дополнялся влиянием на первичную кору полевых сигналов. При отсутствии таковых, прием пищи был достаточным стимулом для чувства насыщения, и связанной с этим удовлетворенностью, но при наличии полевых сигналов чувство насыщения дополнялось соответствующими сигналами полей от товарищей. Игнорировать это невозможно. Если спаривание было не по сезону, полнота чувств все равно требовала контакта с другим. Поскольку и другой находился в том же состоянии, то контакт между ними наблюдатель вполне мог бы трактовать как выяснение отношений, конкуренцию за пищу. Чего греха таить, именно так и трактовал. Есть смысл рассмотреть выражение «полнота чувств» подробнее. Это полное совпадение модели мира, представляемой цели с ощущениями, полученными при ее достижении. Акцептор результата действия получил сенсорный отчет о достижении цели. Внутреннее напряжение, гнавшее к цели, исчезает, наступает релаксация центров и возникает ощущение бесконечной свободы. Возникает ощущение, что энергии избыток, и ее хватит на все, что угодно. Биохимики же говорят что-то об эндорфинах, и диметил-фенил-аминах.
Действия во втором из рассмотренных выше случаев совершенно конкретно были более успешны при наличии стаи. Поле стаи, просто его наличие, было сигналом о том, что помощь близка, удача более чем вероятна, сила страха при этом уменьшалась, появлялся азарт спасения от погони. Когда дело дошло до человека,это явление выразилось в пословице «На миру и смерть красна». В этом случае тот крохотный кусочек коры получал эмоциональное подкрепление, не имея четко поставленной задачи. Кора, даже и древняя, была оптимально активна, не имела четко поставленной задачи, то есть находилась в состоянии оптимальном для свободного творчества. Еще раз напоминаю, что речь в данном случае идет даже не о современных рыбах, а об очень древних их предшественниках.
Третий случай из выше рассмотренных наименее интересен с точки зрения группового поведения. Переваривать пищу и спать вполне можно и одному, при условии, что безопасность обеспечена.
Обонятельный мозг изначально был одним из мест формирования доминанты. Особенностями, отличающими эту доминанту от прочих, было то, что она была наиболее связана с групповым поведением, и здесь формировался интерес личности, выражающийся в поведении.
Рассматривая более развитую кору полушарий, мы видим, что в ней появляется представительство от органов зрения и слуха. Предпосылкой для такого развития было именно групповое поведение, обеспечившее два, казалось бы, несовместимых явления – смертельная опасность, и чувство защищенности в группе. Кора в это время была максимально активна, и для поиска правильного решения (куда бежать) была возможность даже для метода проб и ошибок – не гибла вся стая при охоте хищников. Сигналы в кору от среднего мозга, управляющего бегством, при этом не повергали в бессмысленный ужас, а давали возможность для выработки условных рефлексов. В кору поступали два сигнала – от среднего мозга об опасности, который разрушал личностную доминанту, чтобы она не мешала среднему мозгу действовать – избегать опасности, и от поля стаи, который в это время формировал групповую доминанту. Групповая доминанта при стремлении спастись от опасности была эффективнее индивидуальных среднемозговых доминант. Таким образом, сигналы среднемозговой доминанты все более для коры становились именно сигналами об опасности, о необходимости действовать в группе, а не парализовали кору. Это происходило именно в стае. Среднемозговая доминанта в стае заняла подчиненное положение. Вначале чувство страха, дошедшее до коры от среднего мозга, стало сигналом к включению группового бегства от опасности, а затем зрение и слух перестали информировать кору о среднемозговом анализе ситуации, они сами получили представительство в коре, и уже здесь происходил анализ ситуации, на основе сигналов от рецепторов, не обремененных воплем страха. Первичная обонятельная кора имела множество случайных связей со всеми отделами мозга – таково свойство растущих нейронов.
Таким образом, кора оставила за собой место формирования доминанты, получив от среднего мозга дополнительный информационный канал, точнее два – зрение и слух. Так называемая первично чувствующая кора, более новая по сравнению с обонятельной и вкусовой составила затылочную и височную доли головного мозга. Двигательная кора, тогда же выделившаяся от первичной обонятельной, как и чувствительная, еще долго занимала подчиненное место перед обонятельной древней корой. Обонятельная кора осталась местом формирования главной доминанты личности. Она получила значительное дополнение, входит в состав лимбической системы, и является центром формирования и поддержания эмоций.
Если мы рассмотрим уровень В, то увидим, что структуры, образующие его — зрительный бугор и бледный шар, сформировались в связи с потребностями тела. Зрительный бугор есть часть промежуточного мозга. Здесь происходит афферентный синтез, формирующий картину тела. Интересы поддержания констант гомеостаза лежат в основе работы этого отдела мозга. Реализуются двигательные реакции этого уровня бледным шаром, более молодой структурой, производной древнейшей коры. Подчиненное положение по отношению к обонятельной коре этого отдела мозга обусловлено тем, что на этом уровне основные движения связаны с жизненно важной функцией поддержания гомеостаза у рыб, и с функцией размножения у высших позвоночных. Скажем, рыбы этим уровнем могут еще и локомоции делать, а у высших на этом уровне остались остатки регуляции дыхания, а главным образом этот уровень связан с парным поведением при размножении.
Три важнейших качества этой системы:
1). обширные мышечные синергии;
2). способность столь же стройно и налажено вести движение во времени, обеспечивать правильные чередования, например, перекрестные чередования движений конечностей при локомоциях, объединять в общем ритме;
3). наклонность к штампам, к чеканной повторяемости движений. Все эти особенности движений говорят о способности к длительной непрерывной работе, характерной для вегетативных функций.
Самостоятельно этот уровень ведет движения выразительной мимики, пантомимы и пластики, т.е. совокупность не символических, а непосредственно эмоциональных движений лица, конечностей и тела. Уровень синергий не приспособлен к использованию ни зрительного, ни слухового контроля и управления.
Потому то он и подчинен полностью обонятельной коре и лимбической системе, что без её управления поведением в зависимости от присутствия другой личности, хотя бы и противоположного пола, уровень В как бы хорошо он ни осуществлял синергии, он не имел шансов захватить главенство над уровнем А.
Особенностью уровня Б является то, что он изначально был связан с жизненно важной вегетативной функцией – дыханием. Поэтому этот уровень изначально должен был работать непрерывно. Его непрерывная работа была той базой, на которой строилась его функция. При усложнении опорно-двигательного аппарата с появлением конечностей, конечно же нужны были новые нервные приборы, для управления ими. Этими приборами стала первично-чувствительная кора, как от дистантных рецепторов, так и от тела, причем абстрагированная от рецепции гомеостаза. Так появилась теменная доля. Функции же дыхательного центра перешли в продолговатый мозг.
Сенсорные отделы уровня С расположены в коре головного мозга. Это первичная кора чувствительных зон и ассоциативная кора между ними. Здесь формируется образ пространственного поля. Двигательные зоны этого уровня различны. Уровень С1 имеет двигательный выход от полосатого тела. Этот уровень работает в теснейшем сотрудничестве с уровнем Б, его коррекции идут на базе синергий с учетом ближайших сегментов пространственного поля. Этот уровень (С1) наиболее свободен от влияния лимбической системы, её полевых коррекций, но не свободен полностью. Формируя в движении каждый шаг, коррекции могут происходить с учетом наличия в окружении живых существ, но чаще этот учет происходит в педагогической ситуации, при обучении движениям. Это наиболее консервативный уровень, так как сам является подчиненным, и рано (в онтогенезе) проходит обучение. В то же время так легко подстроить свой шаг под шаг того, кто привлекателен, и очень трудно заставить себя идти в ногу с тем, кто отвратителен.
Уровень С2 производит управление движениями при посредстве пирамидной системы. Нейроны этой системы находятся в задних отделах лобных долей головного мозга, у человека – в прецентральной извилине. Движения этого уровня целевые. Этот уровень развит у рептилий и птиц. Доминанта по прежнему в лимбической системе. В то же время в разных отделах ЦНС появились центры, действующие относительно автономно одновременно.
Появление уровня С привнесло в работу ЦНС нечто новое – в коре головного мозга появились центры, представляющие собой локальные доминанты, действующие полноправно при живой главной доминанте. Эти доминанты, возникающие буквально на каждом шагу, обеспечивают буквально каждый шаг. Афферентный синтез пространственного поля происходит в разных масштабах – для достижения цели оценивается все пространство до нее, каждый шаг – учитывает пространство на протяжении вытянутой руки (ноги). Получается, что главная доминанта раздает свои полномочия для решения частных задач по ходу дела. Локомоции в сложной обстановке требуют разделения конечной цели на ряд промежуточных действий. Взаимоотношения между отделами мозга таковы: лимбическая доминанта держит конечную цель. Акцептор результата действия формулируется в терминах главного в жизни действия. Частные промежуточные цели формулируются под контролем лимбической системы в лобной доле. Задние доли коры формулируют текущее состояние обстановки. Промежуточные акцепторы результата действия расположены в лобной коре. Последовательность их включения определяется в чувствительных зонах, здесь долгое время находился источник сигналов о достижении промежуточных целей. Этот сигнал передавался в лобную долю, вызывая чувство удовлетворения без насыщения, чисто химическим веществом, вырабатываемым здесь же. Этот местный химический сигнал активировал лимбическую систему, вызывая эмоциональное переживание приближения к цели. И на этом уровне в наилучшем эмоциональном состоянии ЦНС была при либо достижении живой цели (у хищников), либо при убегании от опасности. И в том и в другом случае соучастие полевых сигналов способствовало наибольшей активации коры. Получалось, что наиболее активное состояние было в момент напряжения всех физических сил, на пределе возможностей. Здесь то и происходило совершенствование локомоторных актов, и в наилучшем положении оказывались не хищники – направление движения им задавали травоядные жертвы. Бежать они могли в любом направлении, выбор направления и выбор маршрута жертвы задавался не внешней целью, а на базе анализа обстановки, пространственного поля и начального расположения хищника в этом поле. Конечной цели, как таковой, как точки, расположенной в пространственном поле у жертвы не было. Появление хищника вызывало эмоцию страха, вызывало стремление бежать, но выбор направления, маршрута был абсолютно творческим актом. То есть выбор делался свободно, исходя из собственного видения ситуации. Бежать в том или ином направлении, по земле определенной фактуры, менять направление, использовать местные материальные объекты как средства укрытия, все это решалось прямо здесь и сейчас, решение было ответственным, цена ошибки – жизнь. Для жертвы было важно, чтобы решения соответствовали именно этой ситуации, в первый раз новое решение может быть спасительным, во второй раз тот же хищник на ту же удочку не попадется. Решение единственное, годное на один раз требовало в коре свободных ресурсов, и закрепление в памяти именно этого решения было совершенно ошибочным. Ошибочным было и закрепление решения в хромосомной памяти. Хромосомная память была полезной только в том случае, когда закрепляла ощущение постоянной нехватки мозгового ресурса. Наличие свободной оперативной памяти, и памяти на жестком диске определяло возможности наилучшего в данный момент решения. Оперативная память – кора чувствительных отделов мозга. Ее то рост в филогенезе и определяет цефализацию на последних этапах. Жесткий диск – та же кора, но в онтогенезе. Чем больше возможности роста числа и качества нервных клеток и их отростков при развитии конкретного организма, тем больше и разнообразнее его индивидуальный опыт. Незрелость ЦНС на момент рождения, и её созревание в конкретной обстановке делали ее более мобильной. Но если кора обновляется и весь организм обновляется, то как происходит, что личность сохраняется та же? Древняя кора, обонятельная изначально, созревает первой и сохраняется на всю жизнь, доминанта ,созданная на её базе ,ведет нас к витальной цели. Локальные доминанты в других отделах приходят и уходят, а главное остается – еда, вода, дыхание и размножение.
На этом этапе (С) возникло и закрепилось умение строить локальные по месту и времени доминанты, работающие под общим руководством лимбической. Развитие коры при этом преимущественно происходило по линии травоядных. Именно у них первых и пошел безудержный рост чувствительной коры.
Хищники в творчестве были скованы внешней целью, догнать и сожрать – вот и вся философия. Парные взаимоотношения у них тоже были упрощены, сезон спаривания включал врожденные программы размножения, и поводов для личного участия в усовершенствовании поведения не было. Зато их лимбическая система четко дифференцировала полевые сигналы от жертвы и другого хищника. И это тоже был тупик – дифференцировка по принципу «свой-чужой» предельно проста и не требует тонкого различения оттенков настроения. Поскольку уровень С – расцвет гигантских ящеров, то прорыв в млекопитающие осуществили не хищники. Мелкие грызуны, в образе жизни которых было заметно стайное поведение, составили классовую базу млекопитающих.
Итак на уровне С доминанта лимбическая стала передавать доминацию в другие отделы коры. Полной передачи не произошло, все события в коре происходили при доминации лимбической коры. Заслуга ящеров состоит в том, что контроль уровня С1 требовал для коррекции каждого шага локальной доминанты, которая работала ограниченное время, и по совершении положенного включалась следующая доминанта, обеспечивающая следующий шаг. Такая работа тесно связана с уровнем Б, где время составляет чистый ритм. Уровень С2 уже кортикальный, и здесь доминанта задается целью, цель же определяет лимбическая доминанта. В то же время, цель может быть промежуточной, скажем, ящеру надо подняться на гору и обозрев окрестности оценить целесообразность их с точки зрения утилитарной, или попав в яму, не имея перед собой цели – пищи поставить задачу выбраться из нее. Лимбическая же доминанта, будучи устойчивой во времени, санкционировала свыше работу частных доминант, причем главным образом не их последовательное включение, а сам факт их наличия и подчиненного положения. Разрастание чувствительной коры и особенно ассоциативных зон создавало условия для формирования местных очагов возбуждения, мало зависимых от отдаленных участков коры. Эти участки были устойчивы, что и требует условие доминанты, но их устойчивость была ограничена.
Процесс включения доминанты прост: сигнал поступает в кору, и если он требует ответа, в коре происходит распространение возбуждения, до тех пор, пока рефлекс не замыкается и не происходит двигательный ответ – ящер побежал к цели. Но этот процесс протекает по разному у плотоядных и травоядных.
У плотоядных чувство голода активирует лимбическую кору, ее возбуждение генерализуется и пирамидная система сдвигает гору мышц и гонит её за несчастной, ничего не подозревающей жертвой.
У травоядных этот механизм тоже существует, но для генерализации возбуждения в коре, у них есть еще один механизм. Сигнал об опасности от чувствительной коры обладает таким же свойством. В этом случае лимбическая кора не является источником формирования доминанты, хотя она и делает свой вклад. Источником сигналов для формирования доминанты является сенсорная кора. И даже если лимбическая доминанта по старой памяти остается источником устойчивого возбуждения, то сигнал для ее включения лежит вне ее, в другом месте коры. Постепенно, поколение за поколением ведут отбор таких особей, у которых доминанту может сформировать откровенно сенсорная кора. Таким образом у травоядных есть шанс получить принципиальное новшество в коре – способность формировать доминанту не от лимбической коры, но и от сенсорных ее отделов.
Кроме того, стайное поведение давало еще одну важную особенность. Запуск и включение устойчивого очага возбуждения в сенсорной коре сопровождалось изменением работы коры лимбической. Лимбическая кора начинала активно работать именно в направлении усиления коммуникаций с себе подобными. То есть при опасности лимбическая кора не только отказывалась от доминации, но и усиливала свои коммуникационные свойства. В коре возникали две одновременно действующие доминанты, каждая в своем направлении и не гасящие друг друга. Иными словами, доминанта стала структурированной, сложной. Иначе говоря, в коре возник диалог между очагами доминации.
Но и этого мало. При интенсивной физической работе, а убегание от опасности и есть такая работа, от опорно-двигательного аппарата идет массированная импульсация от рецепторов проприоцептивной чувствительности. И еще одним источником импульсов является чувствительность интрацептивная. Собственный организм, информируя кору об изменениях в нем, создает еще один очаг. Таким образом, в коре есть несколько очагов возбуждения при спасении бегством – сигнал об опасности – чувствительная кора дистантной рецепции, сигнал о состоянии стаи – лимбическая кора, сигнал об изменениях в организме – чувствительная кора теменной доли. Все сигналы не подавляют друг друга. Возникает возможность создания функциональных центров с распределенным расположением в коре, а не сконцентрированных в одном месте. Все это вместе взятое потребовало биохимических изменений в тканях организма. Речь идет прежде всего о развитии глиального компонента в нервной ткани, чье развитие позволило локализовать возбуждение, ограничить его генерализацию, без чего формирование нервных центров без четкой локализации, с одновременным участием разных отделов коры было невозможным. Таким образом, развитие нервной ткани и соединительной ткани было уделом травоядных ящеров, каковые и передали это свойство млекопитающим.
Еще одно важное свойство этих ящеров – ограничение в росте. Гормон роста перестал вырабатываться всегда. По достижении определенного размера его выработка прекращалась и возобновлялась только при стрессе. Точнее последовательность была обратной – стрессовая реакция усиливала выброс гормонов, а в стадии восстановления выработка гормонов прекращалась. У хищников дело обстояло наоборот – чувство голода повышало уровень стрессовых гормонов, затем происходило действие, затем утоление голода и прекращение выделения гормонов. У травоядных выделение стрессовых гормонов происходило по сигналу коры, а у плотоядных – по сигналу от рецепторов гомеостаза, и повышение уровня гормонов повышало возбудимость коры. Мы, млекопитающие, произошли от травоядных и должны помнить, что и у рептилий и у млекопитающих двери в будущее открывает травоядность.
Уровень Д.
Вот мы и добрались собственно до неандертальнцев. Именно эти существа довели уровень предметных действий до предельно возможного развития. Современный человек в предметной деятельности далеко превзошел неандертальцев, но и возможности у человека намного больше – он имеет речь.
Уровень предметных действий подразумевает освоение закона причинности вплоть до повседневного его употребления. Выше мы рассмотрели развитие этого уровня. Остается добавить, что лимбическая система и при этом уровне сохранила доминирующее положение. Мало того, у неандертальцев она бесконечно усилилась, по сравнению с австралопитеками. Это произошло вследствие длительной практики совместных действий, связанных с продолжением рода. Дети все более нуждались в обучении, общество обретало все больше разных предметов, а следовательно и умений, связанных с этими предметами. Интенсивность общения все возрастала, а речи все не было.
Сравнивая мозг и голосовой аппарат современного взрослого человека и младенца с тем, что нам известно о древнем человеке, ученые оценивают, насколько человек прямоходящий был способен говорить. Он тоже обладал гортанью, производящей звуки. Но для образования слов звуки должны модулироваться в полостях над гортанью (серый цвет). У современного человека это носовая полость, ротовая полость и глотка. Движение языка меняет форму и объем двух камер — ротовой полости и глотки, — артикулируя основные звуки современной речи.
Как полагают, человек прямоходящий обладал примерно таким голосовым аппаратом, какой изображен здесь и был воссоздан исходя из реконструкций неандертальского человека. Гортань расположена выше, что заметно уменьшает величину находящейся над ней глотки. Относительно длинный и помещающийся почти целиком во рту язык не уходит задней частью в глотку и способен изменять объем только ротовой полости. Такая однокамерная акустическая система делала возможной лишь очень медленную и неуклюжую речь.
Голосовой аппарат современного новорожденного младенца больше напоминает голосовой аппарат человека прямоходящего, чем современного взрослого человека. Язык помещается почти целиком во рту и гортань расположена высоко. Поэтому предполагают, что крик младенца должен напоминать звуки, издававшиеся человеком прямоходящим. Число их невелико, однако мозг взрослого человека прямоходящего мог оформлять их в слова. Но когда мозг современного младенца достигает сравнимой величины, его язык и гортань уже давно успевают опуститься в горле, увеличив глотку и обогатив репертуар его звуков.
Лимбическая доминанта в связи с общением усиливалась, но усиливалось и возникновение локальных, частных доминант, связанных с предметной деятельностью, и локализованных в новой коре. Способности новой коры к созданию и длительному поддержанию доминанты, базирующейся на сигналы из чувствительной коры, то есть от окружающего мира, и имеющей сложную структуру, состоящую из распределенных в коре очагов возбуждения, в противоположность доминанте лимбической, локализованной в одном месте и возникающей по внутреннему побуждению, увеличение площади новой коры, появление и расширение ассоциативных зон, развитие соединительной ткани в организме и глии в нервной системе — все это стало базой для развития предметной деятельности. И приматы, от дриопитеков, освоивших трехмерные локомоции, через рамапитеков, освоивших прямохождение, австралопитеков, потерявших шерсть и освоивших огонь, и, наконец в лице неандертальцев доведшие предметную деятельность до предела возможного на доречевом уровне пронесли знамя антропогенеза до современного человека. Необходимо заметить, что каждый следующий этап в усовершенствовании деятельности происходил скачкообразно при участии климатических катаклизмов. Главным действующим лицом были похолодания. Каждое похолодание стимулировало следующий шаг антропогенеза. В биохимической части работы это рассматривается подробнее. Здесь же следует отметить, что поистине революционные события происходили недолго. Дело в том, что каждое новое умение было прорывом для того, кто сделал его впервые. Сделав этот прорыв при максимальном напряжении всех сил, при генерализованном возбуждении в коре головного мозга, тот, кто впервые сделал защищенное жилье для самки, научил этому свое потомство, наверняка не ожидал, что его потомство попадет в эволюционную ловушку. Суть этой ловушки заключается в том, что если какое то действие становится постоянно повторяемым в нескольких поколениях, то оно легко закрепляется в хромосомном аппарате. Если первый, освоивший теплый дом сделал это в пиковой ситуации, и обученный им сын, в той же пиковой ситуации повторил это действие, то для какого – то количества поколений это будет важным действием, которому обучаются от родителей. Но по мере усвоения действия, условия будут неизбежно улучшаться. И что мы видим? Даже в теплых условиях, при отступлении ледников, или при миграции в теплые края, где необходимости в теплом жилье нет, все равно мы находим следы именно такой деятельности. Австралопитеки два миллиона лет использовали камни определенным образом обработанные, усовершенствование шло крайне медленно, по сравнению с неандертальцами. Дело видимо в том, что с одной стороны, условия были достаточно комфортными, а с другой стороны, в ЦНС происходили изменения, закрепленные в хромосомном аппарате. Как только умение становилось врожденным, прогресс прекращался. Бобры строят жилье, но они строят его не только потому, что нуждаются в нем, но и потому, что у них есть передающиеся по наследству программы строительства жилья, и эти программы включаются независимо от того, есть нужда в жилье или нет. Так же и рамапитеки, я имею в виду ветвь, давшую начало австралопитекам, вели образ жизни, позволивший им сохранить прямохождение не потому, что это им нравилось. Возможно они рады были бы начать ходить на четвереньках, но прямохождение, как врожденная программа была запечатлена в наследственном аппарате, и мешала кое-кому опуститься на четвереньки, хотя такие отщепенцы наверняка были.
Особенно это видно на примере австралопитеков. Множество их видов, обнаруженные археологами, а следовательно образовавшиеся и жившие на протяжении двух миллионов лет говорят нам о том, что локальные условия приводили к такому приспособлению к ним, к такому изменению тела, что это видно на обнаруженных остатках скелетов. К сожалению, о строении их ЦНС очень мало что можно сказать. Мягкие ткани не сохранились. Но увеличение головного мозга и усложнение изготовленных орудий найденных и ныне известных говорит об усложнении их строения в сторону человеческую. Предметная деятельность и ее усложнение и развитие требовали отдельной доминанты по поводу изготовления каждого предмета. Для этого нужно, во-первых, достаточная площадь коры, и во вторых, если доминанта сформирована по поводу достаточно отдаленно похожему на витальный, устойчивая доминанта должна существовать не одна, должна быть иерархия доминант, способная к перестройке и в последовательности включения, и в вариации в силе включения при параллельном включении доминант. Иначе говоря, генерализация возбуждения, необходимая для выполнения действия впервые, должна не только автоматизироваться, с переносом на неосознаваемые уровни управления, но и иметь возможность дозированной генерализации при компиляции уже известных способов действия в необычной обстановке, в необычной последовательности, в необычном сочетании. Постоянное хромосомное закрепление изменений в коре по каждому поводу необходимо было прекратить, а для этого надо было сделать головной мозг максимально незрелым на момент рождения. Чего и добились неандертальцы. Именно они, вид появившийся от вида, жившего в домах, носившего одежду, имевших опыт обучения, а значит и культуру – как способ передачи знаний от поколения к поколению через обучение, — питавшийся обработанной огнем пищей, на протяжении примерно 60 тысяч лет добились того, что обмен веществ в головном мозге стал влиять на хромосомный аппарат. Изменения, полученные в головном мозге искусственным путем, были закреплены в терминах хромосом.
Здесь я имею в виду дугообразный пучок. Пучок волокон, соединяющий у человека зону Брока и зону Вернике в головном мозге, от которого даже следов нет у приматов.
Но об этом позже. Сейчас обращаю внимание на то, что предметная деятельность невозможна без сочетанного действия в трех зонах головного мозга. Если локомоции ящеров требовали возможности формирования доминанты в чувствительной коре, локомоции дриопитеков требовали формирования доминанты в лобной коре, при этом полного подавления лимбической коры не происходило, то предметная деятельность требует полного ее подавления. Я имею в виду неавтоматизированную предметную деятельность, не требующую генерализации возбуждения, т.е. без использования напряжения всех сил организма, но имеющую четко понимаемую смысловую сторону. Дело в том, что при такой деятельности мимика человека имеет специфический вид. Её можно назвать «выражение сосредоточения». В это время ведущей является доминанта, базирующаяся на коре теменной доли и лобной доли. Образ предмета, смысл действия, правильная последовательность действий, и требуют одновременной активности верхней теменной дольки, нижней теменной дольки и премоторной коры. Премоторная кора, начавшая всерьез развитие для усовершенствования локомоций в трехмерном пространстве, в предметной деятельности продолжает развитие. Предметная деятельность, постоянно повторяемая требует развития этой области. Мало того, развитие этой области даже в небольшой степени позволяет формировать нелимбическую доминанту без генерализации возбуждения в коре. Это революционный прорыв. Лобная доминанта позволяет целенаправленную деятельность не только без генерализации возбуждения, но и может подчинить своей власти лимбическую доминанту. Теперь непосредственные потребности тела могут быть отодвинуты на второй план, потом, вот сейчас допишу и чай попью. Кора начинает жить жизнью относительно самостоятельной от наследственных программ.
Значение лимбической доминанты в определении витальных целей остается, но и ее роль в предметной деятельности очень высока. Все обучение, передача знаний от поколения к поколению происходит при непосредственном участии лимбической системы. Отсюда эмоциональное окрашивание всех абсолютно действий неандертальцев, отсюда торможение к дальнейшему их развитию – лимбика требует своего, и, закончив очередной этап деятельности, неандерталец устраивал праздник. Но во время непосредственно предметной деятельности неандерталец был сосредоточен так, как современному человеку вряд ли доступно – усилие воли для подавления лимбики делало неандертальца, преуспевшего в предметной деятельности, йогом.
Эти два наложенных друг на друга черепа (рис. 10) демонстрируют анатомическое разнообразие неандертальцев, подтверждающее возможность того, что они развились в современные типы. Более древний череп (красный цвет) из Ла-Шапель-о-Сен отличается массивностью костной структуры, покатым лбом, срезанным подбородком и выпуклостью на затылке. Наоборот, «Схул V» (оранжевый цвет) с горы Кармель выглядит почти современно — он меньше, имеет четкий подбородочный выступ, высокий лоб и напоминает черепа кроманьонцев, которые пришли на смену неандертальцам.
Умелое переключение с предметной деятельности на чисто лимбическую, при необычайной мощи последней, сделало неандертальца еще и магом.
Еще раз напоминаю, неандерталец не мог говорить, единственное что он мог внятно произнести, это гласные типа А, О, Ы, и губные. Видимо поэтому единственный слог, дошедший до нас как привет от неандертальца – ОМ.
В дошедших до нас трудах Поршнева неандертальцы предстают перед нами как суггесторы небывалой силы. Действительно, общение на доречевом уровне могло происходить только на основе развития эмоциональной сигнальной системы, и требовало постоянных упражнений лимбической системы.
В то же время, на основании многочисленных находок костей животных в местах постоянного обитания неандертальцев находят кости животных, в том числе и очень крупных, на основании чего делается вывод о том, что неандертальцы охотились, и охота для них была одним из основных занятий. Несомненно, охота была очень увлекательным занятием, но с точки зрения развития предметной деятельности она давала очень мало. Охота, как постановка засад, загон в засаду, преследование, выслеживание дичи, постановка её в беспомощное положение с последующим убийством вполне доступна для животных с малоразвитым уровнем Д, и стимулятором развития соответствующих отделов мозга быть не может. То есть неандерталец из племени, которое не занималось охотой впервые приступая к такой деятельности действительно напрягал все свои интеллектуальные способности, и научившись сам, передавал свое умение молодому поколению. Сам он и первые поколения его учеников, несомненно, совершали эволюционный подвиг, делали прорыв, осваивая новую деятельность, но по мере автоматизации новизна уходила и нужда напрягать мозги всерьёз исчезала. Но нам не следует забывать, что развитие предметной деятельности обязательно должно было привести к развитию речи (этот факт невозможно оспорить, если бы было иначе, человечество было бы другим). То есть наиболее прогрессивным способом охоты, с точки зрения неандертальца, стоящего на пути к кроманьонству, был иной способ. Они вполне могли использовать для этого занятия свои суггестивные способности. Выглядела такая охота несколько необычно. Неандерталец-суггестор находил жертву, связывался с ее лимбической системой при посредстве своей лимбической системы, и своей волей, воспитанной поколениями в упражнениях предметной деятельностью, полностью подавлял волю животного, каким бы крупным и опасным оно ни было. После этого животное можно было привести к своему постоянному месту жительства и использовать в качестве пищи. В этом случае неандертальцу не нужны были одомашненные животные, которые требуют определенной заботы и дополнительных трудозатрат.
Еще раз повторяю, что неандерталец совершенствовал предметную деятельность в повседневной жизни, много раз вынужденно повторяя жизненно важные действия, связанные с выращиванием младенцев. Поскольку эта предметная деятельность имела в качестве главного предмета собственного младенца, то параллельно с предметной деятельностью совершенствовались и средства коммуникации. В ЦНС совершенствовались определенные зоны (теменная доля и премоторная кора – для предметной деятельности, и лимбическая система – для коммуникаций). Вся деятельность, связанная с жизнеобеспечением молодых мамаш и их потомства и была тем горнилом, в котором выковывались основы совершенствования человека. Это дом, одежда, тепло очага, питание. Охота суггесторов была вначале забавой, а потом и постоянным занятием.
Но основу для возможностей такой охоты заложили не собственно охотники, а как раз те, кто не мог быть полноценным охотником. Как выше показано, охота хищников в плане развития ЦНС является тупиковым направлением. Вначале это были женщины, совершенствовавшиеся в уходе за младенцами и общении с ними. Это их деятельность была и предметной и межличностным общением одновременно. Совершенствуясь в выхаживании младенцев женщины-неандерталки совершенствовали неандертальцев как вид. С одной стороны, повышение мастерства в выхаживании младенцев не могло не привести к снижению младенческой смертности. А это значит, что по мере совершенствования предметной деятельности, стали выживать и все менее жизнеспособные новорожденные. Причины пониженной жизнеспособности младенцев могут быть разные. Но если парировать незрелость каких то функциональных систем, позволить им созреть в постнатальном периоде онтогенеза, то в следующих поколениях незрелость может стать нормой. В данном случае я имею в виду прежде всего незрелость ЦНС. Из поколения в поколение младенцы неандертальцев становились все более беспомощны, и виновны в этом были сами неандертальцы. Но соответственно, возрастала роль матерей в выращивании потомства, и их умение выхаживать младенцев. Умение выхаживать слабых и немощных, по мере взросления и выхода из детородного возраста, никуда не уходило. Бабушки помогали внучкам, но неожиданно их умение нашло особое применение. Речь идет о больных и раненых. Умение выхаживать привело к появлению первой социальной помощи. Я пишу все это о неандертальцах, это они оставили следы такой помощи больным и раненым.
Говорить о чем-то подобном у австралопитеков не позволяет отсутствие достоверного материала. Таким образом, в группах неандертальцев появились иждивенцы, не способные принимать полноценное участие в предметной деятельности племени. Это говорит о том, что материальная организация племени неандертальцев допускала членство слабых и неспособных к физической работе, иначе говоря, племя имело прибавочный продукт. Речь идет об инстинкте сохранения энергии. Именно неспособные работать физически стали причиной усложнения социальных отношений. Эти неандертальцы, не имея физической занятости на протяжении дня, имели полноценные функциональные системы для предметной деятельности и для общения. Меньше работая физически, они больше работали в направлении коммуникаций. И совершенствовали свои лимбические системы. Последствия этого были удивительными.
Последствия определялись сутью лимбической доминанты в сравнении с доминантой, имеющей в качестве источника другое место в коре. Дело в том, что при всей развитости лимбической системы у неандертальца её свойства базировались на свойствах обонятельного отдела мозга у первых позвоночных. Здесь закладывались структуры, задававшие витальную цель в окружающем мире. Витальные цели – питание и размножение. Гипертрофия лимбической системы у особей, живших на иждивении общества, имела направление коммуникативное, но сам факт гипертрофии структур задававших витальные цели приводил их хозяев к сверхъестественному прагматизму в сочетании со сверхъестественными суггестивными способностями.
Здесь необходимо рассмотреть отдельно мужчин и женщин. Дело в том, что витального значения прагматизм женщин связан с их биологической ролью в обществе. Особи женского пола практически любого вида совершенно неравнодушны к произведениям собственного тела – детям. Лимбическая доминанта делает их поведение достаточно альтруистичным, если речь идет о детях. Жизнь женщины для нее самой может иметь меньшее значение, чем жизнь ребенка, и ответственна за это именно лимбическая доминанта. У женщины она может быть витальна не в отношении особи, а в отношении вида, особь способна на самопожертвование в интересах продолжения жизни вида. Подавление лимбической доминанты у женщины в процессе развития предметной деятельности не заходит слишком далеко.
Иначе дело обстоит у мужчин. Групповое поведение, и прежде всего поведение в процессе продолжения рода так же определяется лимбической доминантой. Но в антропогенезе по мере развития уровня Д у неандертальцев появились возможности временного подавления лимбической доминанты не только при опасности для жизни, но и при простой повседневной деятельности. При условии же, что функции лимбической системы расширяются, все большее значение приобретает ее коммуникативная функция, то природные программы, определяющие половое поведение, и все возрастающую роль мужчины в жизнеобеспечении женщины и потомства могут быть подавлены, и вероятность этого в процессе антропогенеза возрастает. Мужчины слабые физически, но с сильной лимбической системой получили возможность не зависеть от повседневной жизненной деятельности. Их совершенствование пошло в направлении развития как раз лимбической системы. Но коммуникативная сторона обернулась неожиданной стороной. Совершенствование суггестии, сделало их непревзойденными охотниками, они могли обеспечить любое сообщество любым количеством мяса. Но это их умение, будучи примененным к соплеменникам, делало их абсолютными рабовладельцами. А это большой соблазн. Совершенствование в магии и использование обрядов для достижения своих целей, каковые определялись уже не возможностью добыть пищу, не обеспечением безопасности, и даже не борьбой за самку, а на какие цели – никому неведомо. Не было речи и речевое мышление, внутренний диалог или монолог был им недоступен. Зато была доступна картина мира, недоступная современному человеку. С помощью своей лимбической системы неандертальцы-маги видели мир как поток энергии. Им не надо было думать, как достичь той или иной цели – они всегда знали готовый ответ на любой вопрос. Использование магических ритуалов неандертальцами, знакомыми с законом причинности, говорит о том, что все, о чем мечтают современные люди, занимающиеся магией, было доступно многим неандертальцам. Например, в работах Кастанеды говорится, что ключом к магии является остановка внутренней речи. Неандертальцы не знали этой проблемы, у них не было речи. Своей лимбической системой, чувствительность и силу которой они теперь совершенствовали они могли вступать в контакт с себе подобными на огромных расстояниях, но говорили они не долго – не о чем. Будучи всем обеспечены, они не нуждались ни в чем, они были как боги на земле. И главное дело для них было лечение себе подобных, в этом они преуспели так, как нам и не снилось. Главным лечебным воздействием было опять же суггестивное. Закон причинности, постигнутый при помощи структур уровня Д, позволял видеть причинно-следственные связи. Но те же структуры уровня Д, и в первую очередь кора теменной доли и премоторная кора у продвинутых йогов могли формировать устойчивую доминанту, подавлявшую эгоизм лимбической системы. Работа коры совершенного йога, стремящегося добиться гармонии внутренней и внешней позволяла отдельным йогам подняться над личным эгоизмом, и оставаясь созерцателем происходящего, благотворно влиять на него. И такие йоги-неандертальцы становились живыми объектами поклонения, всемогущими богами для соплеменников. А так же и объектами для подражания для подрастающего поколения. Многие хотели научиться жить припеваючи, ничего внешне не делая. Подняться до уровня мага, уравновесить работу лимбической доминанты лобной доминантой, и их совместную работу уравновесить работой теменной доли (созерцание), и все это под руководством ничего не говорящего словами учителя – задача небывалой трудности. Можно подражать внешним действиям при предметной деятельности. Если тебе по лимбическому каналу приходят сигналы одобрения и неодобрения, образные картинки как следует действовать, то созерцать созерцание йога и не заблудиться в его галлюцинациях – задача воистину титаническая. Показать как делать и при этом показать, что надо воздержаться от действия, при этом внимательно следить за извивами бессловесной мысли – это подвиг. Особенно при том, что развитая лимбическая доминанта постоянно требует витальных действий, а ведь она то в этом разговоре учителя и ученика играет ведущую роль.
В любом производстве существуют отходы. Были они и здесь. Не все становились совершенными йогами. Многие, встав на путь совершенствования лимбической системы, быстро попадали в ловушку этой же системы. Она же ответственна за постановку и достижение витальных целей, а ты уже многому научился в плане обеспечения себя материальными благами. Вот они, соплеменники, подчини их себе, и можешь наслаждаться. Чем? Властью. Наслаждаться властью на самом деле невозможно. Власть подразумевает ответственность. Нет ответственности – остается злоупотребление властью. У первых йогов ответственность была обеспечена их физической беспомощностью, зависимостью от материального положения их общества. У их учеников такой зависимости не было. Если они достигали совершенства, то их лобная доминанта уравновешивала лимибическую, а это означает чувство долга, способность перешагнуть через интерес собственного тела. Важным было принять аскетический образ жизни учителя. Он подразумевал служение обществу, которое компенсировало телесные недостатки. Но если телесных недостатков нет, а есть простое чисто животное желание жить по человечески, то обучившись фокусам с лимбическим каналом воздействия на соплеменников, можно позволить себе многое. И это многое и есть злоупотребление властью, в данном случае той, которую дает знание.
Первопроходцы в йоге совершили прорыв и исчезли. Их последователи быстро смекнули, как жить припеваючи на шее соплеменников. Нет, они не были полными бездельниками, они лечили и помогали во всех делах, где знания первых йогов могли пригодиться. Но они ограничили распространение этого знания, стали монополистами и паразитами на теле общества. Речь идет о превышении вреда от них над пользой приносимой ими. Ведь было и то и другое. Но постепенно все больше йогов занимались паразитированием и сделали жизнь неандертальцев невыносимой. Переразвитие лимбической системы привело к тому, что общество неандертальцев стало терять передовые позиции в предметной деятельности. Не прекратили её, а перестали развивать. Простому неандертальцу стало невозможно сконцентрироваться на повседневной предметной деятельности. Лимбический телепатический контроль был воистину полным, всепроникающим и постоянным. Йог-паразит сидел себе на горе и требовал от своего племени… а чего он требовал? Мы этого представить себе не можем – не хватает знаний реалий той жизни. Золотые унитазы – это двадцатый век. У неандертальцев был какой-то аналог золотых унитазов, иначе они жили бы до сих пор, и современный человек бы не возник. Сила действия равна силе противодействия. Вот, противодействуя суггесторам-поработителям, неандертальцы и сделали почти сознательный шаг на пути к человеку. Надо понимать, что и суггесторы и их рабы были одной крови, одного вида. Суггесторы, если и начали выделяться как отдельная раса, делая это путем искусственного отбора, то тем только приблизили свой конец. Дело в том, что рабы суггесторов оказались в положении травоядных перед хищниками. Они должны были научиться молчанию мысли, и подавить полевую связь между собой, и соответственно, отказаться от лимбической коммуникативной системы, а ведь она была в основе системы передачи знаний от поколения к поколению, то есть основой культуры. Отказ от привычного метода коммуникаций привела к поиску новых средств коммуникации. Возможно, это был язык жестов. Тем более действия руками неразрывно связаны с предметной, а следовательно и символической деятельностью. Академик Марр в свое время, высказывал гипотезу о том, что до речи существовал язык жестов. Мы не можем заглянуть в далекое прошлое, не оставившего нужных для проверки этой гипотезы следов. Но в недавнем прошлом был поставлен опыт, косвенно подтверждающий эту гипотезу. Обезьян научили языку жестов, которым говорят глухонемые. Обезьяны сумели освоить от 100 до 300 жестов-знаков, и пользовались ими как при общении с экспериментаторами, так и при общении между собой. Оказалось, что обученные обезьяны явно предпочитали общение между собой или с экспериментаторами общению с другими, не обученными обезьянами. Их группа проявила такую сплоченность, что по окончании эксперимента разделить обезьян представляло большую трудность. Мало того, обучение предметной деятельности у таких обезьян не представляло труда, более того, они сами начинали стремительно учиться и учить друг друга всему, что было на границе обычных обезьяньих возможностей. Мы не знаем, сумели ли неандертальцы-рабы обучиться языку жестов, но если сумели, то лимбическая связь у них явно была уменьшена.
Как ещё защититься от суггестии. Необходимо сосредоточиться на какой-то деятельности, которая бы занимала все внимание, требовала бы максимальной концентрации внимания, и чтобы это внимание было связано с действием собственного тела, и чтобы делать это можно было долго, очень долго. Дело в том, что неандертальцы были субъектами себя осознающими. Это было не массовым явлением, но самосознание было доступно каждому. Ощущение, что все, что ты делаешь, тебе не нужно, ты делаешь потому, что тебе пришел сигнал, принятый твоей лимбической системой как свой собственный, вызывает дискомфорт, особенно если мешает выполнять естественные действия по обслуживанию собственного тела. Этот дискомфорт может быть сильнее или слабее, к нему можно привыкнуть и не замечать. Но земля большая, территории свободные от суггесторов были, и неандертальцы, жившие на этих территориях, прожившие достаточно, чтобы было что вспомнить, вдруг попадали на территорию суггесторов. В этом случае дискомфорт был налицо, привычки к нему не было, как долгое время не было и защиты. Но защита была нужна и ее нашли.
Дело в том, что деятельность не составляющая труда, которой можно заниматься долго и которая будет способствовать самоидентификации проста, как мычание. Это производство звуков. Если ты, находясь под влиянием суггестора-лимбика, начинаешь издавать звуки и звучание тебе нравится, то ты, концентрируя внимание на издавании звука, слушая этот звук, выходишь из под влияния суггестии. Ты поешь и с удовольствием слушаешь свое пение. Твое настроение находится в твоих руках. Как пели неандертальцы?
Голосовой аппарат неандертальцев не позволял им многое. Гласный похожий на Ы и губные согласные – вот и вся песня. Но если концентрировать внимание на звуке ОМ, подобный которому могли произносить неандертальцы, то для противодействия суггестии этого вполне достаточно. Если хватит силы не отвлекаться, а суггестор будет отвлекать, у него собственная гордость. Этот звук вполне пригоден для приведения человека в бездоминантное состояние. Бездоминантное же состояние позволяет четко увидеть, что в твоем поведении совпадает с твоими интересами, а что не совпадает. Чужое внимание становится видимым, вплоть до визуализации источника.
Все это похоже на фантазии. Но есть одно «НО». Я имею в виду дугообразный пучок нервных волокон, соединяющий у человека в головном мозге заднее-нижние отделы лобной доли с верхней височной извилиной. Моторную и премоторную кору со слуховой корой. У человека этот пучок возникает и существует по врожденной программе развития ЦНС, а у прочих приматов его нет, даже у человекообразных.
Так вот борьба неандертальцев с паразитами-суггесторами была важнейшим этапом атропогенеза. Полевой канал коммуникации между личностями исчерпал себя, на смену ему должен был придти другой способ договариваться. Символическая деятельность на базе уровня Д полностью подготовила неандертальцев к созданию речи. Защита от суггесторов заключалась в постоянном повторении звука ОМ с концентрацией внимания на воспроизводимом собственным голосовым аппаратом звуке. Поколения, занимавшиеся этим, так и защищались от суггестии. В результате в хромосомах запечатлелся дугообразный пучок. Он позволяет при обучении речи не только слышать себя, но и совершенно бессознательно контролировать качество произнесения звука, понимать, что издаваемый звук – твое собственное творение.
2.7 КРОМАНЬОНЦЫ
После создания дугообразного пучка дни неандертальцев были сочтены. Оставалось научиться говорить, и стать кроманьонцами. Здесь надо понимать, что возникновение дугообразного пучка вряд ли было сразу распространено. Это произошло на периферии ареала, там, где холодно, там, где имеются дополнительные трудности. Может быть и в Африке, но обязательно там, где холодно, где все описанные события проходили в условиях непрерывного совершенствования предметной деятельности. Падение, уход на боковую и тупиковую ветвь развития происходит там, где это позволяют условия. Взлет возможен тем, кто на самом дне. Лишь тот достоин жизни и свободы, кто каждый день идет за них на бой. Лишь совершенствование методов выхаживания младенцев допустило возможность выходить в семье неандертальцев первого кроманьонца – ведь он наверняка был не самым жизнеспособным из всех. Тем более, за стенами дома – собачий холод.
И вот оно, свершилось. Младенец кричит совершенно неотличимо от остальных. Его голосовой аппарат сразу после рождения не отличается от остальных голосовых аппаратов. Но он растет, и реализация программы его развития неуклонно приводит к закономерному результату – в определенном возрасте он получает возможность воспроизводить звуки, совершенно отличные от звуков, издаваемых его соплеменниками. Но кто научит его говорить? Речь соплеменников назвать речью нельзя. Их общение, построенное на выражении эмоций позой, жестами, мимикой, передачей того же эмоционального состояния полевым каналом лимбической системы это не звуковая речь, и младенец должен научиться говорить сам. Мы знаем, что это невозможно. Примеры с Маугли – человечьими детьми, воспитанными животными — достаточно многочисленны и красноречивы – научиться после пяти лет говорить полноценно невозможно. Речи можно обучиться только от людей и только в определенном возрасте. И здесь мы вступаем в область гипотез, подтвердить или опровергнуть мы не можем. Возможно только два варианта хода событий. Либо говорить научились дети при неговорящих родителях, либо особенностью кроманьонцев была способность освоить речь в любом возрасте, а человек современный именно тем и отличается от кроманьонцев, что его наследственная программа изменилась так, что мозг при рождении максимально незрел, и раннее обучение языку имеет кардинальное преимущество перед самообучением в более зрелом возрасте и мозге способном самостоятельно обучиться голосовой речи.
Мы рассмотрим оба пути.
Прежде всего обратимся к условиям появления кроманьонцев. Итак, неандертальцы распространились по всей земле – Азии, Африке и Европе. Они достаточно приспособлены к жизни практически в любом климатическом поясе. Они могли жить и в холодном климате, питаясь плодами охоты, и в тропическом, питаясь плодами земли, и дополняя рацион той же охотой. Малая зависимость от климата была обеспечена искусственным микроклиматом вокруг тела – дом и одежда уже миллионы лет защищали тела их предков от превратностей погоды. Огонь был им подвластен. Но если их предки – австралопитеки делали дом и одежду при помощи врожденных программ, то неандертальцы делали это при помощи программ, создаваемых по мере надобности. Они имели гипертрофированный уровень Д, и этот уровень был у них высшим, уровня Е, речевого они не имели. Появление и рост каждого следующего поколения требовали умений и знаний по родовспоможению и выхаживанию младенцев, что и определило развитие уровня предметных действий.
Накопленные знания требовали канала связи, по которому их можно было передать следующему поколению. Такой канал связи был создан природой ранее, и неандертальцы использовали его в полной мере, развили и усилили его до предела возможностей. Использование такого канала связи сделало ненужным передачу информации из поколения в поколение по хромосомному, наследственному каналу, позволило каждому следующему поколению рождаться со все менее зрелым мозгом. Под зрелостью я понимаю созревающие в нужное для сохранения вида время готовые к употреблению программы действий. Мозг становился все более универсальным, т.е. не готовым к конкретной работе, но готовым обучиться любой работе. Умение неандертальских женщин выхаживать новорожденных современному человеку невообразимо. У них не было роддомов, памперсов, бутылочек и сосок. В магазинах не продавались предметы женской и младенческой гигиены, да и самих магазинов-то не было. Современная женщина не в состоянии не только самостоятельно родить, она даже и зачать не сумеет, не изучив предварительно подробных наставлений по этому делу, написанных весьма пожилыми академиками, в совершенстве теоретически освоившими предмет настолько, что их слово перевешивает любые природные склонности обучаемого.
Имея лимбический канал коммуникаций, неандертальцы освоили методы суггестии настолько, что мы вправе назвать их магами. Имея прибавочный продукт, вместе с ним общество неандертальцев имело и экологическую нишу для паразитов общества. Паразиты, не имея внутренних ограничений на паразитизм, внешних ограничений не имели тоже, они полностью господствовали над своими соплеменниками, что снижало жизненный потенциал последних. В то же время, неандерталец, подверженный суггестии, не всегда мог отличить собственное желание от наведенного, и вполне мог всю жизнь обслуживать мага, отдавая ему и прибавочный продукт и часть необходимого, и был при этом счастлив оттого, что ему позволено приобщиться священных тайн. Возможно, были местности, в теплых краях, где земля плодородна, где добыть прибавочный продукт легко, и его хватит и паразитам и обществу в целом, там паразитические маги расплодились и имели возможность начать выделяться из общества как особая раса. Возможно, они даже сумели выделиться и как вид, но это маловероятно. Тем не менее, знания магов были огромны, по сравнению со знаниями окружающих их вольных неандертальцев, маги были сильнее мозгом, он у них был конкретно более тренирован. Проблему создавало то, что у магов была чрезвычайно сильна лимбическая система, а она в числе прочего и является тем местом, где возникает желание материальных благ. Именно она и влечет к еде, воде, самке и т.д. Южное сообщество было четко дифференцировано на магов, и их рабов. Когда это стало закрепляться в обычаях, южные маги постепенно теряли квалификацию, оставляя только самые необходимые для сохранения власти знания.
Иначе дело обстояло в местах, где продуктивность почвы была мала, и прибавочный продукт был в небольшом количестве, где был длительный период, когда добыча новой пищи не гарантировалась, и необходимо было иметь запас на голодный период. Само наличие запаса всегда провоцирует появление паразитов, но необходимость противостоять им, была жизненно необходима.
Если в теплых краях жизнеобеспечение могло позволить любые фокусы с паразитированием, то в холодных краях опасность гибели всего племени была если не ограничителем для магов в их паразитизме, то потребностью для остальных защититься от суггестии. Необходимо помнить о том, что помимо паразитирования маги были еще и носителями и передатчиками знаний от поколения к поколению, и их утилитарное значение для выживания вида не вызывало сомнений. Северные неандертальцы в этом отношении не отличались от южных. Маги нужны, но содержание их может быть накладно. Поэтому каждый стремится стать магом. И каждый, не преуспевший в этом, стремился найти защиту от суггестии.
Защитой было:
1. Снизить значение лимбической системы, что в свою очередь требовало создания нового канала связи, что и было сделано в виде второй сигнальной системы;
2. Научиться концентрировать внимание на собственном деле максимально долгое время, жестко противопоставить лимбической доминанте лобную доминанту;
3. Научиться включать и выключать лобную доминанту по своему желанию;
4. Стать независимым от мага, а для этого получить знания магов и не стать при этом паразитом.
Для того, чтобы использовать все эти методы или какой-то из них, неандертальцам нужен был мотив. На севере этот мотив был. На севере все эти методы были использованы неандертальцами. На их стороне был и холодный климат, и очередное наступление ледников. Дело в том, что охлаждение организма у теплокровных для сохранения определенной температуры тела требует активизации работы печени. Повышенный обмен в печени приводит к многим сдвигам в биохимических процессах. Все эти сдвиги описаны в биохимической части работы, здесь же отметим только усиление выработки таурина, который является в числе прочего еще и стабилизатором мембран нейронов, снижает их чувствительность к полевым влияниям, и вполне может использоваться как защита против суггестии. Кроме того, таурин у человеческих новорожденных в больших количествах вырабатывается в мозге, по всей видимости сдерживая созревание не столько нейронов и глии, сколько нервных центров, делая головной мозг в целом менее дифференцированным перед рождением, для того, чтобы подготовить лимбическую систему новорожденного к отсоединению от матери во-первых, и для сдерживания натиска новых впечатлений от внеутробного мира сразу после рождения — во-вторых. Дело в том, что печень во внутриутробном периоде еще не работает в полную силу и не сразу включается после рождения (желтуха новорожденных). Повышенная магическая и противомагическая активность северных неандертальцев и привела к появлению в холодном климате кроманьонцев.
Язык жестов был с одной стороны прямым развитием уровня Д, с другой стороны это был язык, не связанной с речевой системой, дугообразный пучок для использования жестов в качестве средств коммуникации был совершенно не нужен. Скорее всего, этот язык появился у неандертальцев, их развитие это позволяло, но достаточной защиты от суггестии он не давал. Разговор на этом языке требовал предельного усилия лобных долей, что подавляло лимбическую доминанту, но при подавлении лимбики обсуждение эмоциональных состояний становилось невозможным. При обсуждении тонких эмоциональных состояний требовалась дозированная активность лимбической системы, движения рук вызывают эмоцию сосредоточения, и все. Это был язык обсуждения практических действий, конкретных, привязанных к месту и времени, он в достаточной степени тормозил суггестивное воздействие, но не давал возможности по-новому взглянуть на обсуждаемый предмет.
Мне очень трудно себе представить внутреннюю речь, ход мысли, выраженной не в словах, а в рисунке и последовательности жестов. Обдумать в терминах жестов можно только техническую конструкцию, и не очень сложную, обдумать в этих терминах взаимоотношение между людьми мне представляется достаточно сложно. Более того, взаимоотношения между магами и не магами в северной ситуации были намного сложнее, чем в южной.
В то же время органы речи есть одновременно и органы дыхания и приема пищи. Их управление связано с участием лимбической системы, а не с простым ее подавлением. Замена жестов звуками была не просто переключением афферентации с затылочной доли на височную, но это был и переход от предметной символической деятельности при общении, к деятельности символической и эмоциональной одновременно. Лимбическая система не подавлялась, не нарушался эмоциональный настрой во время разговора, переводя все на сухой язык предметной деятельности о предметной деятельности в терминах закона причинности. Появилась возможность говорить о тонких оттенках чувств, эмоций, обсуждать такие глубины восприятия мира, что разговор жестами, если он и был, то быстро забылся. Внесение знаний закона причинности в обсуждение чувств быстро высветлило истинные намерения, избавило человечество (кроманьонство) от необходимости подозрительно относиться к действиям другого, сплотило кроманьонцев.
Здесь мы снова обратимся к опытам советских приматологов. Обезьяны, овладевшие жестовой речью, образовали очень сплоченную группу. Они практически перестали интересоваться жизнью других обезьян. Они предпочитали общение на этом языке, и даже экспериментаторы, представители другого вида, с которыми возможно общение, были предпочтительней представителей своего вида, но не «говорящих». Если говорящие самцы еще могли спариваться с неговорящими самками, то говорящие самки явно предпочитали говорящих самцов, даже вовсе и не доминантных.
Видимо в этом и кроется тайна быстрого исчезновения неандертальцев. Ведь они с кроманьонцами одного вида, но кроманьонцы заговорили, пусть и несовершенно, а неандертальцы – нет. Группа говорящих кроманьонцев сразу стала настолько сплоченной, в ней практически моментально зародился строй, который мы называем «первобытно-общинный» (учившиеся в советской школе понимают, о чем речь). Разница с общественным строем неговорящих неандертальцев заключается в следующем.
Лимбическая система или ее зачатки и по сей день есть у всех видов позвоночных. Она ответственна за общение, и за общественную организацию каждого вида. Общественная организация зиждется на отношениях между особями. Самая сильная связь у млекопитающих – между матерью и детьми. Чем больше дети зависят от матери, тем больше они к ней привязаны. По мере обретения детьми самостоятельности эта связь слабеет. Сильна связь между особями противоположного пола в период спаривания. Это тоже может быть очень сильная связь. Отмечена связь между братьями (сестрами). По крайней мере, многие виды избегают в таком сочетании половой связи. В процессе антропогенеза, по мере отступления наследственной памяти перед индивидуальной, все большее значение приобретает связь между учителем и учеником. Внимательные наблюдатели отмечают в природе и дружеские связи. Все эти виды взаимоотношений есть и в человеческом обществе, они перенесены к нам из животного мира. По мере усложнения человеческого общества, усложняются и связи между людьми в нем.
На самом деле у кроманьонцев по сравнению с неандертальцами мозг имел отличия. Кочеткова отмечает большую величину мозга у кроманьонцев, и резкое увеличение лобной доли. Принимая во внимание, что кроманьонцы овладели и речью, то есть уровнем Е, можно полагать, что в лобной доле преобладание в развитии пришлось на премоторную кору. Скажем, кроманьонцы именно ее развитием и занимались. И здесь возникает самое интересное. Доминанта лобная начинает доминировать и над лимбической и над доминантами чувствительной коры. Особенно это важно для мужчин. Это означает, что у мужчин-кроманьонцев появилось чувство долга, чего уж точно не было у неандертальцев. Чувство долга у мужчин, вплоть до самопожертвования – это и есть способность лобной доминанты подчинить лобной доле все остальные: и эгоистические поползновения — жадность от лимбической доминанты и страх за жизнь — от сенсорной коры. Преобладание лобной коры у кроманьонцев означало и то, что они и словом в карман не лезли, но и действия от них долго ждать не приходилось. Это были кроманьонцы не только слова, но и дела.
Общество кроманьонцев разительно отличалось от общества неандертальцев. У неандертальцев, при их технической вооруженности по сравнению с животными, общественная жизнь не сильно отличалось от таковой у человекообразных обезьян. Лимбическая система у животных в мужских и смешанных стаях заставляет их строить иерархическую лестницу. Волк, вызывающий на бой другого волка, не ведает что творит. Поднявшись над товарищами в стае, он обнаруживает то, чего и предположить не мог, когда дрался за место в иерархии. Пока он был внизу, он был в подчиненном положении, и им помыкали все, кто выше. Взамен он получал защиту от внешнего нападения, участие в более удачной охоте, чем одиночка, а значит и какие-то социальные гарантии. При этом решения принимались не им, вся ответственность за выбор образа действий лежала на вожаках. И вот он на вершине. Но где счастье? Упомянутая ответственность за решения теперь на нем, а все его бывшие товарищи, гарантировавшие ему кусок мяса, пусть и не очень жирный, уступят ему лучший кусок, но все они и радостно закричат «Акела промахнулся», в случае его неудачного решения. Отныне ему спать вполглаза, да слушать вполуха, да опасности ждать не только извне, но и от своих ближайших товарищей. И груз ответственности за каждое решение теперь только на нем. И рабовладение магов-неандертальцев было следствием именно такой стороны действия лимбической доминанты. И несмотря на многие умения, дом, одежду, огонь и основы медицины, да еще и магию, неандертальцы имели общество животных и жили по правилам волков и слонов, особенно мужчины. Самой человечной у неандертальцев была женская часть общества, она и определяла прогрессирование вида на пути очеловечивания. Если у мужчин лучший кусок имел самый сильный (уже не физически, а магически), то у женщин лучший кусок мог получить слабейший, больной, увечный. У неандертальцев наиболее продвинутыми в сторону человека по межличностным отношениям были женщины.
У кроманьонцев наиболее продвинутыми в сторону человека были мужчины. Именно у мужчин лобная доминанта получила максимальное развитие и дала им чувство долга. Кроманьонцы, в силу появления чувства долга, построили общину, которую мы знаем как человеческую. Прежде всего, право силы уступило место праву выгоды для общества в продолжении рода, для семьи а не для отдельной личности.
Наиболее близким историческим примером в описании строя является период военного коммунизма, при большевиках. Но этот пример наиболее известен, менее известно для читателя может быть устройство общины в русских деревнях до социализма. Русские крестьяне в большинстве жили фактически в неолите. Их жизнь мало отличалась от жизни кроманьонцев. У кроманьонцев разве что помещиков не было. Но правила жизни в общине известны, и любопытный может найти их в литературе (Льюис Г. Морган, М.М. Ковалевский, Г.Л Маурэр и т.д.).
Можем заключить, что при преобладании лимбической доминанты общество строится по иерархическому силовому типу, при преобладании доминанты лобной – по типу общины. Именно лобная недостаточность позволяет лимбической системе при организации поведения руководствоваться ставкой на силу и только на силу. Лимбическая доминанта строит поведение, как погоню за материальными благами. Ограничителем здесь является страх наказания и только. Все, что не запрещено, то разрешено – вот лозунг неандертальской лимбики.
Лобная доминанта, как более молодая, не дается при рождении. Она требует длительного обучения. Поэтому мужчины становились полноправными членами общины, когда окружающие оценивали их развитие как достаточное. Сама же система обучения детей с развитой лобной доминантой у кроманьонцев была наивысшей ценностью. Сознательное отношение к общине, основанное на работе лобной доминанты базировалось на связанное с ней чувство долга. Именно это чувство и воспитывалось.
При этом кроманьонец, действуя на любом уровне, должен был руководствоваться лобной доминантой, включать чувство долга, и, при принятии решения, учитывать интересы не просто максимального количества известных ему кроманьонцев, а всех известных ему кроманьонцев, что является очень сложной задачей. Действуя под руководством лимбической доминанты, легче находить решение, главное здесь избежать наказания, а это часто бывает не так уж сложно. Подчинять лимбическую доминанту лобной очень трудно, не менее трудно и учиться этому. Поэтому обществу всегда легче воспитать лимбика, чем лобника. В первобытной общине брак воспитания и давал лимбиков, избежать этого невозможно, отсюда и пошло понятие «мироеды», т.е. люди, научившиеся прямо не нарушать правил общежития, и, тем не менее жить, соблюдая исключительно свои животные интересы. Наказания они избегали, соблюдая минимум приличий. Фактически они в обществе были добровольными изгоями. В то же время, в период, когда небольшое сообщество кроманьонцев было окружено большим количеством неандертальцев, это мало сказывалось на сплоченности кроманьонцев.
Слишком большим наказанием было говорящему оказаться среди неговорящих. Наихудшим наказанием было изгнание из говорящего племени. Таким образом, чувство долга дополнялось пряником в виде возможности говорить, понимать и быть понятым. Отсюда большое количество норм и правил, которые надо соблюдать, потому, что так было решено всем коллективом.
Необходимо сказать, что появление в ЦНС нового уровня управления тонкими движениями, ставшими каналом связи, известным как вторая сигнальная система, было высшим для уровня Д, а согласно Бернштейну, полное развитие любого уровня возможно только при наличии более высокого уровня. Именно этим более высоким уровнем для предметных действий является речь. Появление речи отразилось на изготовлении предметов. На рисунке видно, что изготовление режущих инструментов у кроманьонцев резко отличается по качеству таковых у неандертальцев. То есть требует большего времени, труда, волевых усилий.
Скорее, кроманьонцы учились речи во взрослом состоянии, сознательно осваивая язык звуков, что было для них отчасти продолжением занятий йогой, магией. Именно маги из северных кроманьонцев и создали речь. Солгать было невозможно. Дело в том, что полевой сигнал шел параллельно звуковому и одновременно с ним. Лимбическая система была тренирована на работу с полевыми сигналами, и если при посредстве звуков говорилось одно, а лимбика говорила о другом, то диагноз «ложь» ставился моментально, и это было поводом для выяснения отношений уже без всякой речи – полевой, мимической или звуковой. Женщины, овладевшие речью, явно предпочитали аналогичных мужчин. В общине они получили дополнительные социальные гарантии, чувство защищенности и стали рожать больше девочек.
Передача знаний от поколения к поколению стала иной, чем у неандертальцев. Использование речи значительно облегчило этот процесс и для учителей и для учеников. Причем передача знаний происходила параллельно с овладением речью. То есть ученики учились говорить, и одновременно овладевали знаниями правил поведения в обществе, знаниями прикладными, правилами изготовления и пользования предметами. Учителя же, в том же процессе обучения учились формулировать в словах свои знания, находили новые сочетания звуков для обозначения предметов, этапов их изготовления, правил пользования, правил техники безопасности, и многое другое. Все это делало процесс обучения одновременно процессом усовершенствования жизни. И ученик и учитель, начав обучение, могли не знать, к чему это приведет, они вместе и учились, и учили, и устраивали жизнь, и развивали язык. Все это нашло отражение в речи кроманьонцев. Обучение и развитие речи были неразрывны, становление речи отражалось в становлении общества. Но никакое становление речи и общества не закрепились бы в поведении вида, если бы не имели значения для продолжения рода. Из всех усовершенствований кроманьонцев для продолжения рода имели значение:
а). резкое усовершенствование предметной деятельности на основе возникшей речи, что позволило повысить качество выхаживания младенцев;
б). развитие лобной доли у мужчин и возникновение в связи с этим этической категории долга;
в). возникновение первобытно-общинного общественного строя, с идеологией превосходства рода (вида) над личностью;
г). передача знаний из поколения в поколение с помощью речи позволило расширить запас приобретаемых знаний за счет врожденных. Снова недоношенные дети получили шанс жить, незрелые структуры ЦНС созревали после рождения, и развитие этих структур способствовало становлению именно кроманьонского общественного строя.
Кроманьонский общественный строй имел идеологию. Эта идеология не была врожденной, а следовательно и не передавалась по наследству с хромосомами. Передача идеологии требовала целенаправленного воспитания. В то же время, природные условия на планете не были никогда одинаковы. Более того, они менялись, и жить в разных условиях значило жить по разному. Там, где условия были наиболее трудными, кроманьонцы жили семьями или родами. Первобытно-общинный строй собственно и выковывал волю, характер, и развивал лобные доли, доминирующие над лимбической доминантой.
Там же, где условия жизни были наиболее благоприятными, кроманьонцы могли жить достаточно свободно, не обременяя себя лишними усилиями. Здесь женщины могли жить чисто женскими группами, воспитывая детей, нажитых от случайных встреч с такими же случайными мужчинами. Мужчины же жили в условиях мужских сообществ, не отличающихся от животных стай. Напоминаю, что умение говорить было новинкой, и необязательным для многих умением. Свободные женщины с детьми и свободные мужчины вполне могли создать особое сообщество. Здесь все дети – и мальчики и девочки воспитывались мамами, без пап. Для женщин же формирование лобной доминанты не является обязательным условием для участия в построении здорового общества. Женская лимбическая система сама по себе является гарантией биологически правильного поведения женщины, и как ни развивай лобную доминанту, именно лимбическая заставит её и размножаться, и обеспечит ей чувство долга по отношению к детям. Ошибки в воспитании девочек-кроманьонцев не несли в себе для женского сообщества ничего плохого. Сама природа, в виде требований собственного организма заставляла их обучаться выхаживанию слабых детей, а большего от них и не требовалось. Воспитание же мальчиков требует от них жесткой дисциплины и умения от учителя. Воспитание лобной доминанты процесс сложный, многоступенчатый, требующий все новых знаний, стремления к самосовершенствованию. Всерьёз это могли делать только мужчины. Женщина же воспитывает в мальчике женщину. Мужские наследственные данные воспитываются в женском стиле. Чему могли научить женщины в женских группах неандертальцев? Во первых – потребительскому отношению к мужчинам. Приходящие мужчины должны были сделать то, что недоступно женским силам, и уйти. Женщина, не надеясь на дальнейшую помощь и не рассчитывая на нее, выжимала из пришельца все, что можно, и для этого она использовала свои природные данные, на первом месте среди которых – лимбическая система. Женщины-кроманьонцы не утратили способности к суггестивно-волновым воздействиям, да и у современных женщин они не исчезли. Вот только мальчики кроманьонцы, воспитанные в женском обществе не могли получить лобное воспитание, не реализовывали полностью свой биологический потенциал. Зато они получали наследие предков-неандертальцев. Это наследие не было в первозданном виде. Наследственный аппарат был уже полностью кроманьонский, возможности развития речи на протяжении всей жизни были кроманьонские, но лобная доля нуждается в определенного рода воспитании. А если нет нужного воспитания, то отчетливого, по своей воле установленного доминирования лобной доли над лимбической системой не будет. Лимбическая, как древняя и витальная возьмет свое, и кроманьонец, имея все кроманьонские возможности, включая, упрощенную речь, и связанные с ней усовершенствования предметных действий, в обществе будет действовать как маг-неандерталец. От мамы он научится суггестивно-полевому воздействию на окружающих, он никогда не станет смотреть на общество-общину как высшую ценность и пойдет по тупиковому пути развития своих, к тому времени вымерших предков. Лимбическая доминанта, не обремененная чувством долга, позволит ему выбрать кратчайший путь к витальной цели – еде, воде, женщине, и тому, что позволяет этот путь сократить.
Таким образом, кроманьонское общество было неоднородным, идеология общины и идеология индивидуализма были запрограммированы разными природными условиями, в которых жили кроманьонцы. В силу тех же природных условий кроманьонцы общинники и кроманьонцы лимбики долго не контактировали – жили в разных местах. При встречах, лимбические атаки не могли быть гарантированно удачными, вид кроманьонцев произошел как способ противостояния еще более сильным лимбикам – неандертальцам. Противостоять лимбикам все кроманьонцы могли достаточно эффективно, причем не только на индивидуальном уровне, но и на уровне общества. Правила жизни в общине не позволяли лимбику паразитировать, ничего не отдавая. Лимбик, если он хотел жить в общине, должен был не только выполнять свои общественные обязанности, но и при разговоре не выдать своей склонности к паразитизму, что было не просто – выражение эмоций при разговоре неизбежно выдавало фальшивое поведение – внешнее выражение эмоций и полевое их проявление должны были быть не противоречащими друг другу. Там же где жили одни лимбики, у них не было никаких проблем в общении. Они легко понимали друг друга, и стремясь взобраться на шею друг другу не испытывали враждебных чувств. «Ничего личного» — говорили они, и кто был сильнее тот и садился, а кто слабее – вёз. В основе построения общества у лимбиков было лимбическое поведение, и общество не отличалось от животного – иерархия силы.
Взяв от неандертальцев магию, кроманьонцы лимбики взяли и их образ жизни – максимально приближенный к природе.
Кроманьонцы — лобники строили общество, ставшее основой человеческого общества. И как следствие, живя в более сложных природных и климатических условиях, они, стремясь гарантировать потомство от случайностей, создавали запасы, то есть то, что получали из природы, стремились сохранить на случай, когда из природы взять ничего не получится. В итоге получили прибавочный продукт – приманку для паразитов.
Это одна предпосылка, вторая – по мере роста общины, для её развития неизбежно встал вопрос о разделении труда, и связанный с этим второй вопрос, о передаче знаний, о их концентрации в одном месте, о распоряжении собственностью общины. Вот тут то и вышли на сцену общественной жизни лимбики. Члены общины были связаны родственными узами и наступить на свою лимбическую систему может мужчина, у него лобные доли гипертрофированы, но в отношении своих родственников ему трудно выносить жестокие решения, и, тем более женщина никогда не будет справедлива к кому либо, если он противостоит её детям. Иначе говоря, противоречия внутри общины должен был разрешать кто-то со стороны. Лимбики стали полезны для общины, и увидев это, они стали полезны ещё больше, когда стали не только разрешителями споров, судьями, теми, кто принимает решение, которое может многих не устроить, но не принять которое невозможно. Вот тут их лимбические способности оказались полезны общине. Дело в том, что, живя в условиях трудных, когда биологическая продуктивность почвы мала и нет гарантии иметь средства для жизни на какое-то время вперед, наблюдения за природой многое дают для прогноза и выбора способа действия. Наблюдения за природой естественным образом требуют не только наблюдательности, но и системы запоминания сочетания событий, и системы передачи этих знаний следующим поколениям. И никто не отменял полезности магии во многих случаях жизни. И снова лимбики-кроманьонцы оказываются на службе общества.
Дело в том, что, судя по развитию отделов мозга, кроманьонцы были в значительной степени дубоваты в поведении, эмоции подавляли достаточно эффективно, и жить в обществе кроманьонцев, где никто никого не обманывал, все были, как один, готовы отстаивать свое главное достижение – первобытно-общинный строй, немного скучновато. Скука – недостаток эмоций, функциональный недостаток лимбической системы. Трезвое размышление при помощи заново обретенной речи было намного эффективней жестовой и мимической речи. Она позволила расширить диапазон мышления за счет височной доли, лучше рассчитывать время, и самое главное принимать в расчет эмоциональное состояние свое и партнера. Если эмоциональная привязанность существует и у животных, то у кроманьонцев при составлении пар она не могла не учитываться. Надо сказать, что отношения у неандертальцев мало отличались от отношений у человекообразных обезьян.
Множество авторов, особенно пишущих популярные книги по антропологии, навязывают свое совершенно ничем не обоснованное мнение о половой жизни приматов, навязывая им промискуитет, то есть совершенно беспорядочные связи всех самцов со всеми самками. Более серьёзные исследователи более осторожны в высказываниях. Так в труде М.Ф. Неструх (1970), говорится о том, что человекообразные обезьяны избегают близкородственного скрещивания, что они могут образовывать различные группы – типа гарема, мужские группы, женские с детенышами, смешанные группы, и даже устойчивые на протяжении нескольких лет пары. Что касается кроманьонцев, живущих в теплых краях, то условия им позволяли жить в любом типе сообщества.
Кроманьонцы, живя в домах в условиях холодного климата, скорее жили и семейными парами с детьми, и смешанными группами. Имея тесное общение на протяжении долгой зимы в четырех стенах, явно отбор шел в направлении семейных пар. Лобные доминанты преобладали, и все было поставлено под контроль молодого несовершенного разума. Еще раз оговорюсь, что на земле могли быть любые обстоятельства и любые сообщества кроманьонцев, мы ведем речь только о тех, кто принимал участие в антропогенезе, и вел образ жизни, приведший к появлению современного человека. И появление в совершенно разумной общине кроманьонцев лимбиков, фактически чужаков, позволило усовершенствовать то главное, что продвинуло антропогенез вперед.
Главное, что сделали лимбики кроманьонцы, это вмешательство в составление брачных пар. Дело в том, что не каждому дано видеть человека глазами другого человека. Поводы для ссор на почве любовного треугольника всегда были и будут. В то же время, если пара составлена удачно, то для двоих счастливых ничего больше в жизни не надо. Они, радостно прожив жизнь друг с другом, дадут наилучшее потомство, наилучшим способом воспитанное. И им хорошо, и виду (биологическому) приятно, вид продолжает быть, а чего ему еще надо. Кто-то из лимбиков-кроманьонцев, живущих в общине в качестве мудреца, сделал важное наблюдение, практическое применение которого и создало вид человека современного.
Дело в следующем: правильно составить пару, чтобы эта пара взяла на себя обязательство воспитать члена общества способного поддерживать общественные отношения в таком виде, что общество сохранится, не будет разрушено изнутри.
Одно дело создать общину на пустом месте, когда её ещё не было, когда каждый её член выбирает общество себе подобных из множества других, имея основанием общее коммуникативное свойство – речь. Речь и её носитель будет абсолютной ценностью, не подлежащей сомнениям. Но когда ребенок рождается в таком обществе, где речь используется не в первом поколении, то говорящие окружающие, при полном отсутствии иных, не выделяются ни на каком фоне. Они перестают быть абсолютной ценностью. Вмести с ними утрачивает ценность и общественный строй, даже и первобытно-общинный. Видимо этот общественный строй несколько раз возникал и рушился, ведь дефекты воспитания всегда были, есть и будут.
О правильном составлении пар задумались именно кроманьонцы лимбики. Как лимбики, они чувствовали эмоциональное состояние близких людей, как кроманьонцы они сумели установить причинно-следственные связи между эмоциональными параметрами пар и установили, что наилучшими парами, наиболее устойчивыми, с наилучшим с точки зрения воспитанием детей для общины, являются пары, составленные из кроманьонцев определенных психологических типов. Они выделили четыре категории кроманьонцев по их общественному поведению, внутри которых составление пар можно было проводить без ограничений. До нас дошла эта классификиция в ведической литературе. Там людей, соблюдающих добровольные ограничения в поведении, делят на брахманов, кшатриев, вайшьев и шудр. Эти категории сегодня мы знаем как кастовое деление людей в Индии. Сегодня эти категории людей считаются как профессиональные и общественные группы — касты. У кроманьонцев это были группы брачных контактов. Именно в пределах своей группы следовало выбирать себе наилучшего брачного партнера, с которым будет легко психологически, который по всем психофизиологическим качествам будет создавать вам приятный психологический климат, ведя себя при этом совершенно естественно, то есть не насилуя себя. Сегодня варнашрама-дхарма (так называется классификация людей в ведической литературе – «Шримад-Бхагаватам» ) каждую из означенных групп делит еще на четыре, то есть всего получается 16 групп.
Столько же групп выделяет современная наука, точнее раздел науки – психологии, называется «соционика». В ней так же людей делят на 4 квадры, и в каждой из них выделяют еще 4 группы, всего 16. согласно соционике, наилучшие брачные пары – так называемые «дуальные». Дуальный – значит, что свойства каждого в паре дополнительны свойствам другого. В соционике рассматривают три пары свойств – экстраверсия-интраверсия, логика-этика, сенсорика-интуиция. То есть, например, логико-интуитивному интраверту дуалом будет этико-сенсорный экстраверт.
При беседе со специалистом мы обсудили описанные свойства из классификации варнашрама-дхармы со свойствами соционических групп, и пришли к выводу о их тождественности.
В дуальной паре комфорт каждого обеспечивается тем, что слабые функции партнер прикрывает своими сильными функциями, и ему это ничего не стоит, он ведет себя как хочет. Но это не все. Как правило, ребенок у дуальной пары рождается с качествами одного из родителей. Тогда один из родителей для ребенка является дуалом, а второй – тождествен по психофизиологическим функциям. Люди с одинаковыми соционическими характеристиками видят мир одинаково, они очень хорошо понимают друг друга. Поэтому лучшим учителем является тот, кто вам тождествен. А второй из родителей при этом будет дуалом, то есть человеком, обеспечивающим психологический комфорт. Дети в этом случае получают в лице родителей учителя и доброжелателя. Детям легко и учиться и они вырастают ничего не боящимися, то есть сенсорная кора у них включается без эмоции страха, а значит не надо страх давить, и можно не включать дополнительно лобную долю. Любознательность делает их хорошими учениками. Пример родителей как будущего кажется более чем привлекательным. Стремление жить, как родители, придает обществу стабильность. Бесстрашие не бойца, а познающего, делает такого человека в общении стихийным психоаналитиком. Общение с ним абсолютно благотворно, он не испугается ваших эмоций, он трезво определит, может ли он вам помочь, и честно скажет о своем отношении к вашей проблеме. Если принять во внимание, что все это происходило в общине, где каждый был стихийным коммунистом, и ставил ценности общины выше личных, а значит и при общении друг с другом разговор каждого шел как бы между личностью и общиной, то есть абсолютной ценностью, с богом, а следовательно и предельно уважительно, то несомненно, общение со членом общины было для не членов изрядным потрясением.
Дети дуальных пар вырастали сильными, умными, бесстрашными, внимательными и добрыми людьми. Каждый посторонний, соприкоснувшись с общиной не мог остаться равнодушным, каждый хотел бы быть таким, как члены общины. Безопасность общины долгое время гарантировалась не только редкостью поселений кроманьонцев, но и дружеским к ней отношением со стороны окружающих. Правила поведения в общине стали примером для многих. Если принятие варнашрама-дхармы для неандертальцев было делом их гипертрофированной лобной доминанты (вступать в половую связь не по лимбической симпатии, а по указанию авторитетного лица), то по мере развития общества, значение лобного контроля снижалось, лимбические эмоции все меньше подавлялись и находили себе путь во внешнее выражение, в том числе и при участии речи. Вот тогда то и появились нецензурные слова и выражения. Кроманьонцы поняли, что обсуждение любых вопросов может быть уместно не в любой обстановке. Есть вещи, о которых говорить можно только в ограниченном кругу, иначе их обсуждение может привести к нарушению общественного порядка, не давая ничего для сохранения общества.
Фактически появление мата и стало сигналом о рождении человечества. Это слова, которые вытесняются из речи в человеческом обществе, но могут проявиться внезапно под наркозом, или при операциях на мозге. Составление пар стало фактором искусственного отбора, который и привел к появлению в потомстве кроманьонцев современных людей. Современный человек произошел в первобытной общине во-первых, и в семье во-вторых. Его воспитанием занимались родители и община, под руководством недоразвитого кроманьонца, кроманьонца с признаками неандертальца. Сам он кроманьонцем до конца не стал, но и у кроманьонцев забрал их излишнюю лобную заторможенность, сделал их более эмоциональными, и нашел способ паразитировать на них.
Кроманьонцы имели по сравнению с современным человеком иное строение мозга. Во-первых, мозг у них был больше, во-вторых теменная доля была меньше, и в третьих лобная доля была существенно больше, чем у современного человека. Человек современный по сравнению с кроманьонцем имеем меньшие лобные доли. Скорее всего, они утратили биологический стимул к безудержному росту и воля современного человека в среднем меньше, чем у любого кроманьонца. Но произошли не только количественные изменения. Качественно человек современный не может самостоятельно научиться говорить. Он учится от родителей и только в определенном возрасте. Если до 5 лет человек не имел контакта с людьми, был воспитан животными, то он никогда и не научится говорить, как человек, хотя несколько слов освоить может. Дело в том, что способность говорить была у кроманьонцев. Они могли научиться говорить в любом возрасте. У современного человека такой способности нет. Но у него есть возможность научиться говорить, получить это умение от других в готовом виде. У кроманьонца были структуры, дававшие ему умение создавать речь на ровном месте, и эти структуры были очень энергоемкими, требовали гипертрофированной лобной доли. У нас обучение речи в отсутствие кроманьонских структур требует уже готовых речевых структур, которых нет у нас в том возрасте когда мы учимся говорить, но они есть у наших родителей. Такая утрата произошла потому, что в общине умение говорить у младших активно стимулировалось старшими. Эти старшие и взяли на себя роль утраченных структур.
Поэтому современный человек не может быть как таковой вне всякого общества – без речи он никто.
Внимание к обществу, как внешней среде организма у человека привело к увеличению сенсорной и ассоциативной коры, и в первую очередь теменной доли. Переразвитие теменной доли, где формируется образ предмета и последовательность свершения процессов привела к тому, что человек современный может делать созерцание как основное занятие. Уменьшение лобной доли сделало его менее деятельным, так как общество заинтересовано в том, чтобы человек не столько был деятелен, сколько соблюдал законы. То есть развитие теменной доли тормозит работу лобной доли, а снижение функции лобной доли снимает контроль с лимбической системы. Прорыв лимбической системы к управлению поведением означает животное поведение, совершенно асоциальное.
Если мы примем во внимание, что развитие лобной доли требует целенаправленного воспитания, трудного и для воспитателя и для воспитуемого, что само по себе развитие лобной доли по законам природы автоматически ставит ее под контроль лимбической системы, как определяющей витальные цели, то станет понятно, что сделать человека, способного стать строителем коммунистического общества есть сложнейшая задача. Современные тенденции в образовании стимулируют развитие теменной доли, созерцание как образ жизни, что мы и видим в изобилии в виде наркомании, алкоголизма, компьютерной наркомании, безволии. У всех без исключения насильников, половых извращенцев, у основной массы преступников отмечается лобная недостаточность. Если вы видите человека, который в мечтах совершил много интересного, а в жизни совершенно ничем не проявил себя – это то, о чем идет речь. Накопление знаний без возможности применить их когда либо – и это всё о нем – современном человеке западного типа. Именно там происходит масса немотивированных преступлений, которые есть не что иное, как прорыв лимбической системы к управлению. Теменная доля не может ее контролировать, она подавляет работу лобной доли, а лобная недостаточность и означает пробуждение животного начала.
Обобщая изложенный материал можно утверждать, что в общении у животных между собой и окружающим их миром большое значение имеют различные энергетически – волновые (полевые) формы взаимодействия, в основе которых лежит трансформация энергии химических взаимодействий нейромедиаторов как между собой так и с клетками межнейронных синапсов в электроволновую энергию. В наибольшей степени это характерно для наиболее древних структур мозга.
Так, у дельфинов и китов имеется большой мозг, генерирующий и воспринимающий звуковые, ультразвуковые, инфразвуковые, электромагнитные колебания. Для летучих мышей жизненно важное значение имеет эхолокация, для птиц – ориентация на магнитное поле Земли. И вообще у животных непосредственное ощущение мира через его энергетически — волновые процессы необычайно развито (предчувствие землетрясений, извержений вулканов, цунами и т. д.). Своего максимального развития это ощущение гармонии мира достигло у предшественников человека — неандертальцев, которые, не имея никакого вооружения (клыки, рога, копыта и т.д.), достигли максимальной адаптивности в окружающем их мире и диффузно расселились по всей планете (их останки находят от Португалии до острова Ява) за счет способности к суггестии – неречевого энергетически — волнового воздействия на окружающий мир, прежде всего на хищников и себе подобных но менее суггестивных сородичей (Поршнев Б.Ф. там же). Это был максимум цефализации в мире животных. Однако суггестия порождала защитную реакцию у суггестируемых групп троглодитов — неандертальцев и для этого требовался уже не столько мощный мозг — генератор энергии суггестивного воздействия, сколько мощный фактор контрсуггестивной защиты. Для этого требовалось возникновение в мозге структур, обладающих мощными тормозными свойствами. Эти структуры возникли в виде молодых зон коры серого вещества мозга в лобно – височно — теменной области. Их становление и совершенствование, приведшее к появлению вида человек разумный, сопряжены с непрерывным усилением процессов торможения в мозге, как бы наслоением их друг на друга, с непрерывным усилением контроля за нижележащими отделами мозга. Молодые структуры мозга имеют все более возрастающий метаболический ресурс в виде мощных глиальных структур и интенсивных окислительно – восстановительных процессов в нейронах (А.Р. Лурия «Основы нейропсихологии», 5-ое издание, 2007).
Таким образом, неандертальцы и следующие за ними ископаемые люди кроманьонцы были высшей фазой развития животного мира в плане количественного и функционально- адаптивного совершенствования их мозга — цефализации.
Коммуникативные связи до неандертальцев включительно реализовались на уровне 1 сигнальной системы без включения ассоциативно — мыслительных процессов 2 сигнальной системы. Однако в силу особенностей супермозга неандертальцев их воздействие на окружающих животных было очень мощным и обозначается как суггестия (внушение, психологическое подавление).
Это позволяло последним из троглодитид вести паразитический образ жизни. Они отгоняли хищников от убитых ими крупных животных и питались преимущественно мозгом этих животных, разбивая прочные черепа большими камнями.
Эти факты говорят о том, что между отдельными частями биоценоза существуют мощные коммуникативные взаимосвязи на уровне энергетически — волновых процессов, в основе которых лежит генерация тканями живой материи биополя. Особенно мощным генератором такого биополя является мозг и нервная ткань вообще.
В настоящее время хорошо известно, что в генерации электрофизиологических и других биоэнергетических потенциалов большое значение имеют процессы, протекающие на уровне мембран клеток с участием ряда БАВ (Хазен А.М. « Разум природы и разум человека» М. 2000). Наиболее интенсивны эти процессы именно в нейронах мозга как по образованию медиторов межнейронного взаимодействия так и по интенсивности электрофизиологических процессов на их мембранах (Раевский К.С. «Медиаторные аминокислоты. Нейрофармакологические и нейрохимические аспекты». 1986).
Однако в организме человека и животных имеется и другой важный источник БАВ, участвующих в реализации его защитно — адаптивных процессов- печень. Важно отметить, что печень нарабатывает ряд веществ, необходимых для нормального функционирования мозга и нервной ткани. Это, прежде всего, таурин — вещество необходимое для реализации процессов торможения в нейронах мозга. Благодаря классическим исследованиям отечественных физиологов (Павлов И.П., Ухтомский А.А., Введенский И.Е. и др.) установлено, что наиболее молодые отделы коры мозга человека в области лобно – височно — теменных долей, отвественные за речедвигательную, и во многом, ассоциативно — мыслительную деятельность, сформировались благодаря все более возрастающему усилению тормозных процессов, как бы наслаивающихся друг на друга и создающих контролирующую иерархию высшей нервной деятельности. Такая направленность ее совершенствования у человека не могла быть достигнута без резкого усиления роли печени в адаптивных механизмах и наработки больших количеств универсального тормозного медиатора — таурина, образующегося в печени и перераспределяющегося в различные ткани, прежде всего в мозг. В печени, кроме того, образуются мощные регуляторные вещества, определяющие устойчивость организма к холоду, стимулирующие иммунитет, свертывание крови, заживление ран, а так же устойчивость ко многим другим неблагоприятным физическим факторам.
Здесь необходимо указать, что у животных – млекопитающих печень составляет примерно 4 — 5% от массы тела (а мозг 0,5-1%) и количество таурина у различных животных в тканях различно. Так, у крысы содержание таурина в тканях очень велико, меньшее его количество, например, у лошади или собаки, совсем незначительное количество у морской свинки. И, в зависимости от уровня таурина, в тканях животных их можно разделить на различные по своим адаптивным возможностям по отношению к стресс — воздействиям группы. Выделяют «спринтеров» и «стайеров». Первые имеют большее количество таурина в тканях и более устойчивы к острым и интенсивным повреждающим факторам, а вторые к менее интенсивным, но более длительным. Как крайние варианты, относящиеся к «спринтерам» и «стайерам» можно рассматривать крыс и морских свинок, соответственно, имеются и промежуточные варианты. В наших исследованиях установлено, что эти различия связаны с различной степенью активности образования БАВ в мозге, нервной и соединительной ткани с одной стороны и печени с другой, а также особенностями регуляторного воздействия и обмена их в организме. То есть, условно эти различные адаптивные типы можно разделить на «мозговиков» и «печеночников».
Наши исследования и многолетние наблюдения за здоровыми и больными различными заболеваниями людьми также позволили выделить различные адаптивные типы. При лабораторных исследованиях крови на содержание в ней веществ — адаптогенов, образующихся преимущественно в мозге и нервной ткани, а так же веществ адаптогенов, образующихся преимущественно в печени. Как мы выше отмечали, установлено, что из почти 200 обследуемых у 5% преобладали мозговые адаптогены а у 42% — печеночные. У остальных 53% было различной степени сочетание тех и других.
Это отражалось как на уровне физиологической так и на уровне психологической адаптивности и, соответственно, на особенностях высшей нервной деятельности. Лица с повышенным содержанием таурина и других печеночных БАВ в крови (метионина, аргинина, полиаминов, некоторых липопротеидов и ряда других) были, как правило, выносливы к физическим неблагоприятным факторам (низкая и высокая температура, интенсивная физическая нагрузка, нехватка кислорода, воздействие токсичных веществ, заражение инфекционными заболеваниями и т.д.). Такие люди были более склонны к физическому труду и менее склонны к напряженной умственной деятельности. И наоборот, лица с повышенной концентрацией в крови мозговых БАВ (глутаминовая и аспарагиновая кислоты, цистеин, глутатион, и ряд других) были гораздо менее устойчивы к перечисленным физическим факторам, но гораздо лучше переносили длительный эмоциональный стресс, как правило, являлись людьми творческого умственного труда. Тут уместно дополнить, что первые были в гораздо большей степени склонны к употреблению алкоголя и курению, без каких либо выраженных долговременных отрицательных последствий для их организма, что объясняется гораздо более мощной системой детоксикации в их печени и большей стабильностью их мембран в тканях за счет стабилизирующего воздействия таурина.
Поэтому лабилизация мембран алкоголем и защита от интенсивного обмена эндогенных БАВ вводимыми при курении БАВ — ксенобиотиками, прежде всего в печени, оказывает антистрессовое воздействие в условиях хронического, например, эмоционального стресса, воздействующего на людей с преобладанием «печеночного» адаптивного типа.
Напротив, на людей «мозгового типа» курение и алкоголь оказывают резко отрицательное воздействие, приводя к быстрому развитию различных серьезных заболеваний.
Эти выводы согласуются с результатами отечественных исследователей (К.В. Судаков К.В. «Церебральные механизмы эмоционального стресса» 2004, Менделевич В.Д.» Неврозология и психосоматическая медицина» 2002 и др..), а также зарубежными авторами (У. Матурана и Ф.Варела- сантьягская школа когнитивизма), отмечающими, что физиологические механизмы адаптации тесно связаны с механизмами адаптации психологической на уровне регуляции высшей нервной деятельности медиаторами — веществами, выполняющими в организме адаптивные функции. И познание окружающего мира – это есть также своеобразный процесс адаптации, но уже не только отдельного организма, а популяции или общества в целом, адекватно приспосабливающихся к изменениям окружающей действительности. Здесь уместно также привести понятие о культуре как форме популяционной адаптации (Ю.И. Новоженов «Филитическая эволюция человека». 2005).
Эти представления соответствуют концепциям великих отечественных физиологов И.П. Павлова, А.К. Ухтомского, И.Е. Введенского и др. о ведущей роли центральной нервной системы в адаптивных механизмах организма. Эти идеи развивались такими видными и разносторонними учеными как Б.Ф. Поршнев, Л.С. Выготский, пришедших к заключению о том, что физиологические и психологические механизмы приспособления к изменяющимся условиям среды изменяются как у отдельного индивидуума в процессе его развития а затем старения; так и у отдельных популяций (стоящих на разных ступенях социально — исторического процесса) в целом.
Для человека и человеческого общества важнейшим фактором адаптивности является формирование виртуального образа — программы, позволяющей стремиться к определенной намеченной цели, используя при этом весь накопленный опыт. Адекватность виртуального образа окружающим условиям, его пластичность и динамичность в соответствии с изменениями этих условий, во многом определяются психофизиологическим потенциалом индивидуума и общества, интенсивностью реализации этого потенциала.
При этом происходит одновременное развитие многих аспектов психологической адаптивности представителей развивающегося народа: языка, сознания, мышления, эмоций, чувств и т.д. (Поршнев Б.Ф., Выготский Л.С. Марр Н.Я.).
Согласно нашей концепции при этом происходит изменение соотношений психофизиологических типов от преобладания «печеночников» к «мозговикам».
Мы уточняем эти положения тем, что выявили различные адаптивные типы, зависящие от особенностей стресс — воздействий среды и социального окружения человека. В крайних вариантах это «печеночники» и «мозговики».
Установление факта, что существуют различные адаптивные типы и как крайние их формы «мозговые» и «печеночные» позволяет предположить, что непосредственные предшественники человека неандертальцы были в большей степени «мозговиками» а окружающие их млекопитающие «печеночниками». Между ними происходил интенсивный биоэнергетический обмен: суггестивное воздействие троглодитов на животных и, видимо, получение от них «печеночной» энергии, что стабилизировало биоэнергетику биоценоза в целом, гармонизировало его с окружающим миром. Максимальная цефализация, которой достигли троглодиты – суггесторы, означала тот факт, что они, а затем и их потомки — люди, заняли доминирующее положение в плане влияния на биополе биоценоза.
Но условия на планете непрерывно изменялись: исчезали хищники, поставляющие троглодитам крупных животных для питания, наступал ледник с резким снижением температуры. Рассеянные по планете суггесторы — потребители неандертальцы резко утратили адаптивность и стали быстро вымирать. Выжили лишь те ветви, которые смогли относительно быстро по геобиоцинотическим меркам перестроиться на преобладание печеночных типов адаптации. Это позволило резко повысить устойчивость к экстремальным условиям. Стали быстро формироваться структуры мозга, ответственные за координацию движений, средства коммуникаций – восприятие и воспроизведение речевых сигналов, что возможно было лишь в условиях резкого повышения роли печени в адаптивных механизмах и повышения устойчивости к токсичным веществам, низким температурам и другим экстремальным факторам. Сложились реальные предпосылки становления речи, соответственно, мышления, а, значит, и человека как такового. Первичной формой человеческого бытия явилась первобытная община, для которой в фазе дикости характерно полное растворение индивидуума в коллективе, психологическое подчинение всех общинников образу — дипластии, переводящему фантастические представления в реальную действительность, являющегося ее стержнем.
Возникла небывалая до тех пор в природе форма адаптивности – социальная адаптивность, основанная на образном восприятии мира и коллективном вхождении в этот образ, например, все считают себя потомками собаки или медведя и т.д. (тотемизм). Что представляло собою единый организм, противостоящий окружающей его природе, другим общинам и сохранявшимся еще троглодитам — суггесторам, то есть, носило, прежде всего, антисуггестивный характер. Это уже была другая коммуникативная энергетика нежели у троглодитов – неандертальцев – энергетика, генерируемая коллективом — общиной. «Дикари» — общинники представляли собою уже полный антипод не способным создавать устойчивые группы суггесторам — неандертальцам. Это были уже ловкие и сильные охотники, всем своим существом преданные общине, воспринимающие мир как некое агрессивное окружение, живущие в постоянном страхе. Центром такой общины среди ее общинников — «печеночников» был вождь, колдун или шаман — суггестор, по каким то тонким природным механизмам воспроизводивший генетические и психологические свойства своих предшественников «мозговиков», воспринимающих гармонию мира непосредственно, на медитативном уровне. Но это были уже не потребители — суггесторы. Они выполняют роль материализованного образа — дипластии и являются носителями духовного стержня общины.
3. ПСИХОФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ СОЦИАЛЬНОЙ АДАПТИВНОСТИ
3.1. ДИНАМИЗМ СОЦИАЛЬНО-ЭНЕРГИТИЧЕСКИХ УРОВНЕЙ ОБЩЕСТВА.
Таким образом, в первобытной общине появляется новая форма адаптации — социальная адаптация, которая являет собою некую энергетическую сущность, довлеющую над коллективом и дающую опору для психологической адаптации в экстремальных окружающих условиях а это, в свою очередь, резко повышает и физиологическую адаптивность каждого отдельного индивидуума. Важнейшим механизмом возникновения такой социально — энергетической сущности является взаимодействие между «печеночниками» — общинниками и «мозговиками» — суггесторами — вождями, колдунами и шаманами, находящимися в ее центре, а затем другими представителями элиты общества и его основания. В монографии Поршнева Б.Ф. мы находим, что проводились соответствующие расчеты (М. Рубнер), показывающие, что энергетические ресурсы, поступающие с пищей у человека затрачиваются в основном на адаптационно – коммуникативные процессы (то есть психологические адаптивные механизмы) а у животных — на обновление структур организма, терморегуляцию и другие физиологические адаптивные функции. Различие в 5 — 6 раз.
Важно отметить, что особенности физиологической и психологической адаптивности дикарей первобытной общины соответствуют дипластивному стилю мышления, то есть, восприятию реальности с одной стороны как существующей действительности, а с другой как фантастических не связанных с реальностью образов (Л. Леви – Брюль 1930). Такое мышление характерно для детей раннего возраста и связано с особенностями обменных процессов в мозге, прежде всего с содержанием в его ткани больших количеств таурина. Повышенные концентрации таурина отмечаются и в мозге детенышей животных (Раевский К.С. там же). Однако таурин у детенышей животных и у современных детей образуется преимущественно в самой ткани мозга, как и многие другие адаптогены организма, позволяющие пережить экстремальные условия родов, переход на дыхание атмосферным воздухом, первый контакт с микрофлорой и т.д. У взрослого человека таурин нарабатывается в основном печенью и перераспределяется по всему организму преимущественно в мозг и нервную ткань, для чего существуют специальные транспортные каналы. Этот факт еще раз подтверждает правомочность выделения «печеночного» психофизиологического адаптивного типа, характерного для дикарей — общинников и в той или иной мере сохранившегося до наших дней у народов, живущих в естестве первичной общины. Однако гео-климатические, этно-демографические, социально-исторические процессы отличаются неравномерностью. У отдельных народов окружающие их условия претерпевали резкие изменения, и для того, чтобы выжить в этих условиях, необходимо было к ним приспособиться. Изменившийся характер стресс-воздействий требовал и изменения психофизиологической адаптивности индивидуума, а соответственно, и социальной адаптивности популяции, общества. Основываясь на работах Г.Л. Маурера, Л.Г. Моргана, М.М. Ковалевского, Ф.Энгельса мы можем проследить социальную трансформацию человеческого общества от естества первобытной общины фазы дикости к фазе варварства, затем к зарождению первых рабовладельческих государств и империй, становлению феодального, а затем и капиталистического общества. В процессе чего первобытная община трансформировалась в соседскую, затем сельскую, а потом все больше и больше разрушалась, пока не исчезала совсем у народов, достигших высот цивилизационного развития (Эллада, Рим, Западноевропейские государства, США).
Из имеющихся в нашем распоряжении источников мы знаем, что зарождение человеческой цивилизации происходило примерно в 5 — 4 тысячелетии до н.э. Первичные очаги этой цивилизации были локальными и практически друг с другом не связанными (Шумер, Хараппа, Микены, Хаттусас, Угарит, Египет древнего царства, древний Китай и т.д.).
Однако из имеющихся источников ясно, что это были цивилизации непосредственно трансформировавшиеся из естества первобытной общины с приматом жреческой патриархальности, мощными внутренними связями крестьян-общинников, имеющих дипластивное мироощущение (преобладание «печеночного» типа) и относительно чистых «мозговиков»- жрецов, определявших все ключевые стороны жизни общества того времени. В этом плане интересно отметить, что имеющиеся у нас скульптурные портреты представителей жречества и высшей знати обществ 3 — 2 тыс. лет до.н.э. (например Эхнатон, Нефертити и их дочери) имеют форму головы сильно вытянутую назад и гораздо большую чем у современных людей. Если учесть, что похожую форму головы имели троглодиты-суггесторы и ископаемые люди кроманьонцы (Поршнев Б.Ф. там же), то очевидна непосредственная генетическая преемственность высшей знати и жречества от «мозговиков» — троглодитов (а затем колдунов, шаманов и патриархальной верхушки доисторического общинного мира) и их способности мощного восприятия гармонии окружающего мира и трансформации ее в суггестию окружающих их общинников — «печеночников». Эти же обстоятельства могут объяснять некоторые феноменальные способности жрецов древнего мира, например, их астрономические знания, технологии возведения мегалитических построек и ряд других труднообъяснимых, а порою и недоступных современной науке знаний, что дает поводы к спекуляциям на тему о контактах с внеземными цивилизациями. И, разумеется, в таком обществе невозможно было рабовладельчество, ибо его резкая поляризация на небольшую группу жрецов, знати и окружающую их крестьянскую массу и так подразумевало безраздельное адапто-психологическое подчинение ее правящему классу. Причем подчинение это было добровольным, исходящим из дипластичного мировосприятия дикаря и затем и крестьянина- общинника, эмоциональное удовлетворение которого состояло в том что бы отдавать все (даже свою жизнь) в пользу коллектива а не забирать из него, как у предшествовавших индивидуалистов- потребителей троглодитов – суггесторов (Поршнев Б.Ф. «Социальная психология и история» 1979.)
Такая трансформация стала возможной благодаря тому, что в защитно — адаптивных механизмах общинника на уровне его мозга и нервной системы вообще преобладают уже не процессы возбуждения и суггестии, а процессы торможения и мощного коммуникативного защитно-адаптивного взаимодействия внутри своей общины. Такая психологическая защита в доисторические времена была направлена как против чужих общин (ОНИ И МЫ) так и против сохранившихся троглодитов — суггесторов. И противостояние это было необходимо как мощный стимул совершенствования адапто-психологической защиты (Поршнев Б.Ф. там же).
Существуют многочисленные примеры, что изолированные по тем или иным причинам племена могут тысячелетиями не претерпевать процессов социального развития, оставаясь в фазе дикости, не имея даже своего вождя или колдуна (Дж. Фрезер «Золотая ветвь» 2001).
Великий испанский философ Х. Ортега-и-Гассет отмечал («Восстание масс» 1937, «Бесхребетная Испания» 1921.), что народы, обладающие мощными жизненными силами и ступающие из естества природы в цивилизацию как субъекты исторического развития, выдвигают из своей среды лучших. Эти немногие заряжаются духовной энергией массы, которая фокусирует на них свою духовную энергию, но одновременно способна безоговорочно повиноваться им и тем самым творить из них Элиту, высшую творчески-созидательную касту общества. Именно элиты инициируют созидание государств и империй, развивающих цивилизацию. Для других народов, не достигших необходимого уровня социального развития и не имеющих необходимого потенциала жизненной силы, цивилизация является скорее ядом способным убить их, как, например, североамериканских индейцев.
Таким образом, для того чтобы происходило интенсивное развитие великих народов по пути цивилизационного развития — необходима их внутренняя структурализация, основывающаяся на определенной социальной гармонии. Наиболее полноценная для цивилизационного развития общественная структурализация ( по мнению Х. Ортеги – и – Гассета -там же )имела место у германских народов в эпоху их феодального развития.
Однако по мере становления и развития государств и империй, а затем достижения ими зенита и упадка в обществе все более нарастают процессы обособления групп, классов, индивидуумов. Общественная гармония все более трансформируется в противостояние: патриархальная верхушка, колдуны и шаманы и общинная масса, а затем жречество и знать и крестьянская масса первых цивилизаций. Рабочий класс и буржуазия государств нового времени. Со временем нарастает напряжение как между общественными группами и классами внутри цивилизационных обществ, так и между ними в виде восстаний, войн, революций и т.д. Это создает социально – энергетическое напряжение, стимулирующее психологическую адаптивность общества, на базе которой формируется культура как форма его адаптации (Новоженов Ю.И. «Филитическая эволюция человека» 2005). Или, другими словами, совершенствуется его социальная адаптивность как некий энергетический феномен, существующий над обществом в целом в виде его социально — энергетической иерархии и, соответственно, над каждым отдельным индивидуумом соответственно его общественному статусу. Это мощно стимулирует, развивает его психику и мышление. Социальная психология в форме энергетически — полевого напряжения базируется на психофизиологической адаптивности народа в целом, который представляет собою некий гибрид из соотношений «мозговых» и «печеночных» адаптивных типов.
Соотношения эти непрерывно меняются в соответствии с изменениями окружающего мира и, соответственно, изменяются социально – энергетические уровни общества.
В определенных точках развития возникают «золотые сечения» — оптимальное соответствие социально — энергетического потенциала популяции предъявляемым цивилизационным стресс нагрузкам. Процесс этого развития имеет свой пик в виде духовного единства наиболее активных представителей народа, когда он способен на многие казалось бы иррациональные вещи: религиозное одухотворение, героизм, самопожертвование во имя дающей энергетический потенциал доктрины — то есть, к максимальному духовно-энергетическому единству, как психологическому состоянию характеризуемому в социальной психологии как сериация. Именно этот период соответствует молодости народа, открывающей перспективы цивилизационного его развития. В психологически — мировоззренческом плане сериативный тип мышления следует за дипластивным, когда накопление определенного количества дипластий – образов существенно расширяет кругозор индивидуума и социума, но они еще не могут выбирать из них оптимальные для себя программы. При этом доминируют стиль, мода, подражание, еще сильны иррационалистические духовные ориентиры – религия, поклонение святыням и т.д.
Именно в этот период возникает наиболее мощное социально — энергетическое поле, максимально взаимодействующее с биополем биоценоза и окружающего мира, что дает энергетическую основу озарений – восприятия гармонии мира и путей гармонизации с ним в данный цивилизационный период. Озарения эти являются чистым «мозговикам», подпитывающихся энергетикой окружающего преимущественно «печеночного» общества. На этой основе затем формируется цивилизационная культура как форма адаптации, как это было, например, в Эпоху Возрождения у народов Западной Европы. Где рыцарски – романтическое одухотворение « золотого века» феодализма 11 — 12 веков стало в дальнейшем энергетически – социальной основой этой эпохи.
В историческом аспекте все более нарастающие цивилизационные стресс нагрузки требуют включения в процесс цивилизационного развития все более многочисленных и адаптивно более выносливых народов.
Здесь следует также указать, что в психофизиологическом плане помимо изменений в популяции соотношений «мозговики» — «печеночники» изменение цивилизационной адаптивности народа сопряжено с появлением промежуточных форм, в которых, конечно может преобладать либо печеночный, либо мозговой тип адаптации. Интересно отметить, что промежуточные, более сбалансированные формы более характерны для женщин, тогда как для мужчин более типичны крайности, что определяется физиологическими особенностями мужского и женского организма. При этом если мужчина более ориентирован на реализацию себя как личности в социо — культурном развитии общества, то для женщины на первом месте стоит семья, она преимущественно оценивает потенции мужчины в этом обществе, его статус секс (Новоженов Ю.И. «Статус секс и эволюция человека» 1991). Эти различия способствуют усилению социально — энергетического напряжения в обществе, стимулирует процессы его психофизиологической адаптивности. И, напротив, в эмансипированных, чрезмерно усредненных психофизиологически обществах это напряжение спадает и соответственно нарушается адекватная стресс — нагрузкам стимуляция адаптивных ресурсов — нарастают различные психологические расстройства как социальные, так и отдельных индивидуумов. Именно такой процесс психофизиологической деградации в настоящее время мы и наблюдаем в некоторых экономически развитых странах, на первый взгляд вполне благополучных, с их демографическим спадом, широким распространением феминизма и гомосексуализма, гедонизма и бездуховности.
Таким образом, только выход социальной адаптивности на уровень постоянной и интенсивной трансформации адаптивных типов и дает право говорить о начале процесса цивилизационного развития у того или иного народа и составляющих этот народ индивидуумов. В процесс развития по этому пути включаются все новые и новые более многочисленные народы. При этом крестьянская община непрерывно сокращается, а социальная надстройка общества нарастает. Изменяется характер и интенсивность духовно- энергетического напряжения в обществе при существенном видоизменении его социальной адаптивности. При этом, несмотря на то, что полярность «мозговики» — «печеночники» ослабевает, в целом социально — энергетическое напряжение возрастает. Что происходит благодаря все большему включению в цивилизационный процесс все более и более многочисленных народов и способствует его ускорению.
Как установлено Поршневым Б.Ф., трансформация эта происходила на фоне изменений психологических особенностей человека и окружающего его социума («Социальная психология и история» 1979). Формирование дипластивного типа мышления у дикаря – общинника, сменяется сериативным мышлением у представителя элиты жреческого или военно- политического общества, и наконец, в условиях развитой культуры, например, капиталистического общества доминирует уже дихотомичный тип мышления, подразумевающий выбор наиболее оптимальных образов- программ, которыми руководствуется общество и большая часть его индивидуумов.
С нашей точки зрения, расслоение первичной общины и ее дальнейшее разрушение в ходе вступления народов на цивилизационный путь развития был сопряжен с изменением соотношений «мозговики» — «печеночники» в сторону снижения доли «печеночников» и возрастания количества «мозговиков». И вообще снижения уровня «печеночности» отдельных индивидуумов. В силу все более меняющегося характера стресс нагрузок (уменьшение воздействия экстремальных физических факторов и нарастания эмоционально — коммуникативных — «социальный стресс»). Однако интенсивно увеличивающаяся численность народов вступающих в процессы исторического развития позволяла создавать социальные духовно — энергетические уровни адаптивности адекватные современным цивилизационным стресс — нагрузкам и постоянно повышать свою адаптивность.
Для каждой эпохи пик социально — энергетической адаптивности был различным и качество этого пика характеризовалось тем «золотым сечением» выстраивавшейся иерархии общества, которое в максимальной степени соответствовало современным тому времени стресс нагрузкам. Здесь необходимо остановиться на замечательной книге Шривишнупада «Манифест варнашрамы» 1993 года, в которой указывается, что согласно ведической традиции любое общество, развивающееся по цивилизационному пути, имеет свою внутреннюю духовно — энергетическую иерархию, носителями которой на ее верхнем уровне являются мыслители — брахманы, ниже стоят воины — кшатрии, обеспечивающие волевые военно- политические функции государства, затем идут вайшьи — торговцы, ремесленники и другие представители общества, обеспечивающие его материальные потребности и, наконец, на нижней ступени стоят шудры — слуги всех трех вышеозначенных групп.
Мы еще можем добавить, что пятой группой является крестьянская или кочевническая община, ибо общинники живут в совершенно иной системе стресс — воздействий, нежели представители любого уровня социальной надстройки. Как отмечается Шривишнупадой, все перечисленные иерархические уровни общества есть не кастовое его деление, а духовно — энергетические уровни, создаваемые генетически предрасположенными к этому людьми. Кастовое деление возникает и костенеет уже в периоды деградации и упадка прошедших свой жизненный путь народов. И только те общества способны достигать высот цивилизационного развития и внутренней гармонии, где имеются мощные брахманический и кшатрийский уровни, то есть там, где сильны дух, мысль и воля.
Аналогичные идеи высказывает Х. Ортега — и – Гассет (там же) о том, что кастовость общества в духовно – энергетическом плане опирается прежде всего на аристократически – кшатрийское мировоззрение властителей. При этом каждый народ, реализуя свои жизненные потенции, создает особенные узнаваемые духовные культуры. Так, феодализм лежал в основе многосторонней творческой активности народов Европы эпохи Возрождения, и затем предпринимательского творчества ее капиталистических государств- империй. Интересно отметить, что философ приходит к выводу о том, что как его родная Испания, так и в еще большей степени Россия к началу ХХ столетия не прошли этапа феодально — кшатрийской духовной структурализации общества. Располагая колоссальным жизненным потенциалом они непременно должны пройти этот этап как прелюдию подлинного социально – исторического развития.
Мы считаем, что градация общества на социально — энергетические уровни во многом обусловлена соотношением в нем психофизиологических адаптивных типов и их качественными изменениями. Для древнего мира и первых локальных цивилизаций это был жреческо — брахманический тип с небольшой группой «мозговиков» жрецов в центре социально — энергетического поля того общества и большой массой крестьян — «печеночников» вокруг. Сама по себе локальность цивилизаций древнейшего мира и порою поражающий величием уровень их культуры свидетельствует о мощной энергетической подпитке «мозговиков» за счет «печеночников». Следующий этап – кшатрийские военно- политические общества — империи. Спектр высших слоев общества существенно расширяется за счет трансформации бывших крестьян- общинников в кшатрии. То есть, это уже не чисто «печеночники», жившие в естестве природы, а люди смешанного психофизиологического адаптивного типа — «печеночники»- «мозговики», но с явным преобладанием печеночного компонента, способные переносить большие физические трудности и обладающие высоким адаптивно- волевым потенциалом, физиологически базирующемся на мощных тормозных процессах «печеночников». Однако освобождение из под тормозного влияния печени мозгового адаптивного компонента и значительное увеличение количества людей, его носителей, в связи с непрерывным ростом населения предопределяет изменение качества духовно — энергетического уровня такого общества по сравнению с предшествующим – брахманическим. Этот уровень несравненно более мощный и открывает его мыслителям в виде озарений все новые принципы сакральной геометрии природы, о которых говорили еще пифагорейцы (некие пропорции, законы окружающего нас мира, постижение которых приходит к нам в виде озарений). Именно кшатрийские государства заложили основы современного мировоззрения и способов постижения окружающей нас действительности: древняя Индия, Китай, Ассирия, Персия, Эллада, Рим.
3.2 ТРАНСФОРМАЦИЯ ПСИХОФИЗИОЛОГИЧЕСКИХ АДАПТИВНЫХ ТИПОВ КАК СОЦИАЛЬНО — ЭНЕРГЕТИЧЕСКАЯ ОСНОВА ИСТОРИЧЕСКОГО ПРОЦЕССА
Суть изменений состоит, прежде всего, в том, что на первых этапах (фаза дикости) усиление тормозных процессов блокирует и ставит под полный контроль максимально развившееся у предков – предшественников (троглодитов) мозговое суггестивное «ОНО», основанное на процессах возбуждения и генерации подавляющих все живое биоэнергетических волн. При этом формируется первый социальнопсихологический организм – «МЫ», противостоящий окружающему миру: сохранившимся троглодитам – суггесторам, другим таким же МЫ, хищникам, стихийным силам природы и т.д. В таком организме отсутствует осознание его членами «Я», ибо оно невозможно по объективным, экстремальным условиям существования, выжить в которых возможно только сообща, растворившись в едином социальном организме, имея мощные механизмы контроля над необузданным мозговым реликтовым «ОНО». Это уровень дипластивного мышления, когда взрослые члены общины живут в мире сказки — фантазии, безоговорочно веря, в магическую силу колдуна или шамана а их дети, находясь постоянно в экстремальных условиях существования, гораздо прагматичнее взрослых, ибо еще не могут в полной мере приобщиться к этой сказке, прочно цементирующей общину как единый социальный организм. Это обстоятельство как и жертвенность во имя общины ее адаптивно- виртуального образа (принесение людей в жертву, членовредительство и т.д.) есть атрибут полного подчинения адаптивному образу-программе и соответствующее эмоциональное удовлетворение в этом, что возможно лишь при полном блокировании какого либо «Я» (Поршнев Б.Ф. 1974., Л. Леви Брюль 1930). Это же подтверждает пуналуальная семья — многомужество и многоженство у диких народов, которая невозможна при ревности и чувстве первенства у членов общины. Эти факты удивляли многих исследователей и не находили рационального объяснения.
Существенное повышение адаптивности человеческих сообществ по сравнению с окружающим животным миром способствовало разрастанию общин и последующему их расслоению, накоплению знаний, опыта, что приводило к выделению патриархальной верхушки общины и, соответственно, к все более отчетливому осознанию «Я» ее представителями. Как мы уже отмечали, дальнейшие процессы расслоения и разложения общины лежали в основе зарождения первых государств при существенном изменении конфигурации виртуального образа-программы, лежащей в основе адаптивности данной популяции. У представителей определенной части общины происходит понижение «печеночного» потенциала с разблокированием потенциала «мозгового». Реликтовое «ОНО» которых могло в определенной степени удовлетворять свои потребности, отличные от потребностей остальной общины.
Находясь включенными в социально- психологический развивающийся организм представители верхушки были вынуждены жить по законам его развития, испытывая на себе психологическое воздействие окружающих преимущественно «печеночных» членов. Однако все большее количество «Я» создает некую новую социально- психологическую общность, которая ощущает свое «МЫ» уже в своей замкнутой группе, зачастую противостоящей окружающим общинникам — «печеночникам». Характерно, что жертвенность по отношению к виртуальному образу общины трансформируется у представителей надстройки в готовность жертвовать собою ради виртуального образа этой надстройки. Так весьма опасны ритуалы инициации у жрецов древнего мира, часто заканчивающикся гибелью или увечием рыцарские турниры — ритуалы у феодалов Европы, допустимость гибели на поединке от руки человека своего круга, жизнь как подготовка к смерти у японских самураев, харакири при «потере лица» — то есть выпадение из виртуального образа – все это проявления «Я» как психо-адаптивного феномена надстройки общества.
Таким образом, жертвенность первобытной общины во имя самой общины трансформируется в жертвенность во имя виртуального образа надстройки. Разблокирование реликтового «ОНО» и трансформация его в «Я» в условиях развития социального организма вело к формированию нового «МЫ» — надстройки общины, подпитывающейся энергией «печеночников». Все это резко повышает энергетический потенциал такого общества и способствует его структурализации. Виртуальный образ- программа становится все более и более персонализированным, ведет к структурализации общества и повышению его адаптивности. Безоговорочное непосредственное поклонение смутно представляемым силам природы (тотемизм, анимализм, шаманизм) трансформируются в монотеистические религии.
Соприкосновение с «НИМ» высшим началом — Богом, опосредуется представителями жрецов, затем церкви и орденов феодальных сословий, через короля, царя, императора как помазанника божьего. При этом происходит все большее включение «мозгового» компонента в психоэнергетику общества, способствующего все более глубоким озарениям и постижениям гармонии окружающего мира. В недрах церкви возникает наука как новый все более адаптивный виртуальный образ этого мира. Так духовно — энергетический иррационализм и фанатизм религий (этап преобладания сериативного мышления) трансформируется в детерминистское научное мировоззрение. Особенностью доминирующего мышления этого периода является дихотомия, с преобладанием «мозгового» индивидуального и социального типа адаптивности , то есть выбор наиболее рациональных для каждого индивидуума виртуально — адаптивных образов. На этой основе формируется наука, законодательство и другие формальные институты общества.
3.3 ВОЗМОЖНАЯ РОЛЬ И МЕСТО РОССИИ В СОВРЕМЕННОЙ ЦИВИЛИЗАЦИИ.
История России имеет свои особенности. В период раннего средневековья Россия развивалась вровень, а то и опережая ведущие западно-европейские государства, где ее называли Гардарика страна городов. Киев в домонгольский период был больше чем Лондон и Париж. Дочь Ярослава Мудрого была выдана замуж за французского короля, невесткой Владимира Мономаха была англо-саксонская принцесса и эти примеры можно продолжать очень долго. Основой процветания феодального государства был торговый путь «из варяг в греки», однако в силу того, что к Хlll веку он во многом утратил своё значение, экономика и государственность Руси пришли в упадок (Б. Когарлицкий. 2004). Над ней нависла угроза иноземного вторжения, которая реализовалась в «татаро-монгольском иге». В Х111 – Х1У веке государство пережило еще 14 мощных вторжений кочевников, до основания подорвавших его социально – экономическое устройство. Крестьяне и жители городов бежали в труднодоступные для монгольской конницы местности. Опустели старые центры Киевской Руси.
Но время шло, и вместо старых центров стали образовываться новые. Наибольшее значение в становлении нового русского государства имело Московское княжество. Бывшая окраина Киевской Руси постепенно превращается в центр Московского государства. Именно иго обусловило ускоренное, не по внутренним потенциям, создание централизованного государства, и Московская Русь Ивана 111 во многом стала преемницей Золотой Оpды (Трубецкой Н.С., Савицкий П.Н., Гумилев Л.Н. «Россия между Европой и Азией»» 1983). В результате слишком ранней централизации власти Русь была избавлена от межфеодальных войн, что, в свою очередь, привело к замедлению развития, в отличие от Западной Европы, военной техники и других технологий. Это привело в результате к огромному технологическому и научному отставанию, ибо общеизвестно, что прогресс науки и технологии во все времена определялся заказами военных. Жизненно важное преодоление таких отставаний осуществлялось, как правило, по инициативе сверху, рывками, за счет беспощадной эксплуатации труда крепостных крестьян, на чьих костях возводились города, заводы, крепилась мощь армии, а подлинного развития общества не было.
При этом важно отметить, что помимо изменения политических и экономических центров государства произошел своеобразный откат в социально историческом плане. На смену достаточно развитым феодальным отношениям в государственном устройстве пришло доминирование патриархальной, доисторической общины. Феодальная вольница была резко ограничена. Князья и бояре Московской Руси это уже не удельные князья Руси Киевской и тем более не бароны и графы Западной Европы. Они стали такими же слугами (холопами) царя, как и крестьяне общинники. Опираясь на последних, на простонародье, московские великие князья, а затем цари вели жестокую борьбу с феодалами (например, опричнина) а заодно и с феодальным развитием государства. Антифеодальную направленность носила деятельность Дмитрия Донского, Ивана III, Ивана IУ, Петра I. Русский дворянин находился на службе у царя, и на кормление ему давали поместье и крестьян. Но их могли и отобрать, дворянина могли судить за ненадлежащее обращение с ввереной ему землей и крестьянами. Служивый дворянин (а служить должны были все мужчины дворянского происхождения) должен был находиться на военной службе вплоть до неспособности по возрасту выполнять эту службу. Только тогда он получал отставку. Так продолжалось вплоть до екатерининского времени, то есть до крнца ХУIII века. И только в ХIХ веке русское дворянство постепенно распоясалось вкупе с появившимися и бурно увеличивающимися в числе купечеством и буржуазией. Однако основной опорой государства по — прежнему оставалась крестьянская община.
Крестьяне были организованы в сельские общины, которые решали все жизненные вопросы, волновавшие крестьянина, вплоть до того, что если барин становился нестерпимым для общины она могла выбрать человека, который убивал своего хозяина, и потом вся община содержала выборного на каторге, а по возвращении встречала как героя ( Мухин Ю.И. 1993).
В силу этих причин Россия к началу ХХ века оставалась во многом «доисторическим» общинным государством. По переписи 1916 года в стране было 85% крестьян общинников, то есть ещё сто лет назад в России преобладал «печёночный» адаптивный тип.
Культура, сформировавшаяся в России к началу ХХ века, носила в основном дипластивный характер, отражая мировоззрение основной части общинного населения, откуда идут духовность, соборность, патриархальность и прочие атрибуты «исконно-русской культуры». Глубокая пропасть между доисторической общиной и прозападной элитой явилась основной причиной социалистической (общинной) революции, вернувшей все на круги своя в Российской своеобразной общинно – циклической цивилизации.
Последовавшие затем урбанизация и индустриализация, проведенные Сталиным, мало чем отличались по характеру от деятельности предшествовавших ему правителей. Это был возврат к преобладанию общинных форм общественного устройства над феодально – капиталистическими.
Но в ходе этих преобразований к 50 – 60г.г. ХХ столетия крестьянская община в России истаяла. Городское население стало превышать сельское, а к концу столетия крестьян осталось лишь несколько процентов.
Все мы во 2 -3 колене из крестьян, ибо аристократическая и буржуазная элита общества была либо уничтожена, либо выслана из страны. Но проблема в том, что столь быстрое разрушение крестьянской общины (за десятилетия) не позволило сформироваться истинной элите общества. У пассионарных народов Запада и Востока эти процессы занимали столетия. За это время успевала сформироваться и приобрести необходимую преемственность элита пассионарного народа и основанного им государства. Особенно выраженную преемственность вплоть до наших дней такая элита, как уже отмечалось, имела на Западе.
В настоящее время уровень развития нашего общества таков, что мы можем рассчитывать только на переход от разрушенного дипластивного общинного уровня культуры к сериативному, характерному для аристократически – кшатриской фазы культуры как формы адаптации. За советский период, в процессе индустриализации и урбанизации была практически полностью разрушена крестьянская община и её духовные ориентиры. Сейчас у нас отсутствует нормальная структуризация общества в силу того, что не создана соответствующая нашим потенциям сериативная культура. И подлинная интеллигенция, брахманы нашего общества должны помочь ее становлению, путем формирования более нравственной и ответственной его элиты. Что предсказывал нам еще в 1927 г. один из выдающихся мыслителей и интеллигентов России Алексеев Н.Н.
Приведенные нами аргументы о психо – физиологическом (вплоть до метаболического уровня) и социально – энергетическом состоянии нашего общества позволяют утверждать, в противовес мнениям о нашей необратимой социальной деградации, что оно имеет высокий потенциал развития от преобладания «печеночного» компонента к компоненту «мозговому», что позволит создать новый виртуальный образ – программу, существенно повышающий его адаптивность в современном мире. В дальнейшем это позволит нам выйти на новые уровни науки, искусства, философии и другие стороны духовного развития с формированием принципиально нового мировоззрения, соответствующего стрессовым нагрузкам современного мира.
Это единственно возможный путь преодоления системного кризиса, поразившего человечество и грозящего скорой глобальной катастрофой. Все здоровые силы человечества должны присоединиться к нам. Только так мы можем создать ИНТЕГРИЮ – новую форму культуры как форму адаптации к колоссальным вызовам современности, ибо империи с их угнетением человека человеком как инструмент прогресса и форма общественной адаптации себя полностью исчерпали.
ЗАКЛЮЧЕНИЕ
Проведенный нами анализ основ жизнедеятельности и развития живой материи с момента ее зарождения до современного состояния биоценоза с позиций различных научных подходов позволяет утверждать, что основным свойством и одновременно механизмом ее совершенствования является адаптация- приспособление к изменяющимся условиям окружающей среды. При этом, если на ранних этапах зарождения живых существ их совершенствование происходило на уровне обменных процессов , то затем адаптивные механизмы все больше проявляются на уровне функциональном-повышается их активность и плодовитость. По мере остывания планеты возникла необходимость в использовании биоценозом энергии Солнца, что было достигнуто формированием процессов фотосинтеза некоторыми бактериями и растениями. Однако при этом, как побочный продукт фотосинтеза в атмосфере стал накапливаться кислород — мощный окислитель, который произвел революцию в развитии живой материи. Наиболее приспособленной к использованию кислорода тканью является нервная ткань и особенно головной мозг. Именно их возникновение и совершенствование во многом определило дальнейший процесс эволюции (процесс цефализации — постоянного увеличения и совершенствования мозга). На этом пути наибольшего совершенства достигли млекопитающие, среди которых максимум цефализации приходится на предшественников человека-троглодитов — неандертальцев, которые, как теперь установлено, людьми еще не были. Но их огромный мозг позволял им адаптироваться за счет генерации особого рода энергии — суггестии (внушения, подавления) окружающего живого мира и жить как потребители, отбирая добычу у хищников и своих менее суггестивных сородичей, а затем стать их пожирателями-каннибалами. Однако эти суггестируемые вынуждены были вырабатывать механизмы контрсуггестии — мощные тормозные процессы с участием активного совершенствования механизмов адаптации на уровне печени, контролирующих функцию мозга. Так возникли люди — общинники, мозг которых стал меньше по размеру но в нем появились структуры, ответственные за речь и мышление и высокую координацию движений. С этого момента появляется и все более совершенствуется механизм психологической адаптации — в основе которого противостояние суггесторов и контрсуггесторов, дающее мощную энергетику биополя, подавляющую другие генерируемые участниками биоценоза энергии.
По мере развития человеческого общества (троглодиты неандертальцы вымерли в силу утраты адаптивности) это противостояние по генетическим и социально энергетическим механизмам трансформируется в духовно — энергетическое поле. Это поле интегрирует социальные группы общества и создает определеную их иерархию с противостоянием классов и социальных групп, формирующихся при распаде естества первичной (крестьянской или кочевнической общины) народов, еще не ступавших в полной мере в процесс цивилизационного развития.
Духовно — энергетическое поле дает основу для развития культуры как формы адаптации. При этом постоянно совершенствуются психофизиологические механизмы с участием мозга и печени. Россия, недавно вышедшая по историческим меркам из преобладания естества крестьянской общины, имеет высокий потенциал социально – исторического развития.
ЛИТЕРАТУРА
1. Абашева Г.К. Свободные аминокислоты в плазме крови и головного мозга у детей с синдромом Дауна // Журн. невропатол. и психиатр. им. Корсакова. –1981. –Т.81. –Вып. 7. –С.1068-1070.
2. Абашева Г.К. Свободные аминокислоты плазмы крови и головного мозга у детей с синдромом Дауна // Хим.-фарм. журнал. – 1991. — № 2. – с. 17-19
3. Анисимов В.А. Влияние геропротекторов на канцерогенез // Успехи совр. биологии. – 1991. – 92. – 3. – с. 455-472
4. Анохин П.К. Биология и нейрофизиология условного рефлекса. – М.: Наука, 1968. – 548 с.
5. Арчаков И.И. Микросомальное окисление. — М.: Наука, 1975. – 327 с.
6. Афанасьев И.Б. Свободно-радикальные ингибиторы и промоторы в биологических процессах // Кислородные радикалы в химии, биологии, медицине: Сб. тр. — Рига, 1988. -С. 9-25.
7. Белова Т.И., Судаков К.В. Морфофункциональные изменения мозга в условиях эмоционального стресса // Вестн. АМН СССР. – 1990. — № 2
8. Беляев Н.А. и др. Влияние этанола на кефалино-окоидную систему мозга крыс // Биохимия 1984. – Т.49. – вып. 9. — с. 1425-1430
9. Березов Т.Т. Обмен аминокислот в опухолях. – М.: Медицина, 1969. – 195 с.
10. Березов Т.Т. Коровкин Б.Ф. Биологическая химия. / Учебник под ред. акад. АМН СССР С.С.Дебова. – 1990. – 528 с.
11. Бернштейн Н.А. Физиология движений и активность. – М.: Наука, 1990. – 496 с.
12. Бехтерева Н.П. Мозговая организация эмоций // Вестн. АМН СССР 1975. — № 8. – с. 8-19
13. Бехтерева Н.П. Мозговая организация эмоций и активность симпатоадреналовой системы у обезьян в условиях острого и хронического эмоционального стресса // Изв. АН СССР. – Сер. биолог. – 1985. – № 6. – с.822-831.
14. Большакова Т.Д. и др. Активность симпато-адреналовой системы у обезьян в условиях острого и хронического эмоционального стресса. // Изв. АН СССР. Сер. биол. – 1985. — № 6. – с. 822-831
15. Буров В.Ю., Ведерникова Н.Н. Нейрохимия и фармокология алкоголизма. М.: Медицина. – 1985. – 238 с.
16. Вальдман А.В. и др. Психофармокология эмоций. М.: Медицина 1976. – 124 с.
17. Вальдман А.В. и др. Фармокологическая регуляция эмоционального стресса. М.: Медицина. – 1979. – 360 с.
18. Вернадский В.И. Биосфера и ноосфера. / Предисловие Р.К. Баландина. – М.: Айрис-Пресс. 2004. – 576 с. (Библиотека истории и культуры).
19. Волков М.С., Генкин А.Н., Глотов Н.А., Маевский Е.И. Глутаминовая кислота. Биохимическое обоснование практического использования. – Свердловск, 1975. — 119 с.
20. Герштейн Л.М. Содержание белков в нейронах мозга крыс предрасположенных и устойчивых к эмоциональному стрессу // Бюлл. эксперим. биол. и медицины. 1998. — № 10. – с. 477-480
21. Голиков П.П. Рецепторные механизмы глюкокортикоидного эффекта. -М.: Медицина, 1988. — 288 с.
22. Голиков С.И., Саноцкий И.В., Тиунов Л.А. Общие механизмы токсического действия. — М.: Медицина, 1986. — 278 с.
23. Горбунова А.В., Каштанов С.И. Биогенные амины в ганглиях автономной нервной системы при эмоциональном стрессе // Физиол. журн. СССР им. Сеченова. – 1990. – т. 76. — № 2. – с. 176-183
24. Градусов Ю.Н. Усвояемость аминокислот. – М.: «Колос». – 1979. – 400 с.
25. Гуляева Н.В., Левина О.л. Действие алкоголя при хроническом эмоционально-болевом стрессе // Бюлл. эксперим. биол. и медицины. – 1985. – т. 100. — № 8. – с. 204-206
26. Гумилев Л.Н. От Руси до России. Очерки этнической истории Кер. Вехи истории. СПб. Кос. Изд. дом «Кристалл». 2002. – 352 с.
27. Гумилев Л.Н. Этногенез и биосфера Земли. Ин-т Дн-Дик. 1997. – 640 с.
28. Джиоев Ф.К., Плисс Г.Б. Практические и научные основы профилактики канцерогенных воздействий. – Л. – 1984. – 160 с.
29. Западнюк В.И., Куприн Л.П., Занка Л.У., Безверхая И.С. Аминокислоты в медицине. — Киев.: Здоров’я, 1982. — С. 58-63.
30. Иди М. Недостоющее звено. Пер. с англ. И. Гуровой. Ред. а предисл. Ю.Г. Рычкова. М., «Мир», 1977. – 166 с.
31. Казначеев В.П. Проблемы адаптации и конституции человека на севере// Бюлл. Сиб. Отд. АМНСССР.-№1 С.95-99.
32. Капра Фритьоф. Скрытые связи / перев. с англ. М.: ООО издательский дом «София». – 2004. – 336 с.
33. Караедова Л.М., Горенштейн Б.И., Нефедов Л.И., Климович И.И. К ме¬
ханизму гепатопротекторного действия таурина// Мед. новости.-1998. — №9. — с.54-55.
34. Караедова Л.М., Нефедов Л.И., Климович И.И. Формирование пула таурина в условиях стимуляции синтеза НАД+ // Фармакологические свой¬ства новых химических соединений и некоторых лекарственных препаратов. Материалы конференции.-Минск,1994. — с.100-101.
35. Ковалёв И.Е. Действие 20-метилхолантрена на антителообразование // Вопросы онкологии. — 1969. — № 9. — с. 72.
36. Ковалёв И.Е. Иммунитет как функция системы организма, инактивирующей чужеродные химические соединения // Хим.-фарм. журн. — 1977. — № 12. — С. 3.
37. Ковалёв И.Е., Полевая О.Ю. Антитела к физиологически активным соединениям. — М.: Медицина, 1984. — 126 с.
38. Ковалевский М.М. Общинное землепользование, причины, ход и последствия его разложения. М. 1879
39. Ковалевский М.М. Первобытное право. М. 1876
40. Когарлицкий Б. Периферийная империя. –М.: Ультракультура. –2004. –528.
41. Коматов Е.А. Нейромедиотарная интеграция эмоционального возбуждения и механизма устойчивости к стрессу // Вестн. Росс. АН. 1995. — № 11. – с. 9-16
42. Констэбл Дж. Неандертальцы. Пер. с англ. И. Гуровой. Ред. и предисл. Ю.Г. Рычкова. М., «Мир», 1978. – 159 с.
43. Кричевская А.А., Шугалей В.С., Цветненко Е.З. Аргиназа и полиамины мозга и печени в механизме защитного действия аргинина при гипероксии // Бюлл. экспер. биол. и мед. — 1981. — Т.91, № 4. — С. 445-447.
44. Кузнецов С.А. Клетки иммунной системы как посредники в реакции других систем организма на стресс воздействие // Патологическая физиология и экспериментальная терапия. – 1997. — № 2. – с. 27-29
45. Лабори А. Регуляция обменных процессов // Пер. с фр. М.: Медицина. – 1970. – 176 с.
46. Леви-Брюль Л. Первобытное мышление // Изд. «Атеист», 1930.
47. Линдауэр Р. и др. Нейробиология посттравматического стресс расстройства // Соц. и клиническая психиатрия. – 2003. — № 1. – с. 446-450
48. Линдауэр Р. Нейробиология посттравматического стресс-расстройства // Соц. и клинич. психиатрия. -№ 1. –С.464-150.
49. Лукьянова Л.Д., Валмуханов Б.С., Уголев А.Т. Кислородзависимые процессы в клетке и её функциональное состояние. — М.: Наука, 1982. — 301 с.
50. Лурия А.Р. Основы нейропсихологии: учеб. пособие для студ. высш. учеб. заведений // А.Р. Лурия – 5-е изд., стер. – М.: Издательский центр «Академия», 2006. – 384 с.
51. Манифест Варнашрамы. Практическое руководство по реорганизации общества Харикенша Свами по заказу партии Санкиртана // Изд-во «Бхастиведанта Бук Траст». г. Екатеринбург. 1993. – 260 с.
52. Матурана Умберто и Варела Франсиско. Древо познания. –М.:Издательство Прогресс Традиция. 2001г. – 224 с.
53. Мдзинарашвили А.М. и др. Влияние вальпроевой кислоты на структуры сна и потребление этанола у крыс с различными типами индивидуальной реактивности до и после стресс воздействия // Бюлл эксперим. биологии и медицины. – 1989. – т. 108. — № 9. – с. 294-296
54. Меерзон Ф.З., Малышев И.Ю. Феномен адаптационной стабилизации структур и защита сердца. М.: Наука. – 1993. – 157 с.
55. Меерсон Ф.З. Защитные эффекты адаптации и некоторые перспективы развития адаптационной медицины // Успехи физиол. наук. –1991. –Т.22. -№2. –С.52-89.
56. Меерсон Ф.З., Малышев И.Ю. Феномен адаптационной стабилизации структур и защита сердца. – М.: Наука. –1993. –157 с.
57. Менделевич В.Д., Соловьева С.Л. Неврозология и психосоматическая медицина. – М.: Мед-пресс. информ. 2002. – 608 с.
58. Моисеенко В.С., Загорулько Л.И., Тадор И.П. Особенности кислородного обмена при опухолевом процессе // Журн. эксперим. онкология. – 1985. -№ 5. –Т.7. –С.8-14.
59. Морган Л.Г. Древнее общество. – Л.: Изд. Института народов севера ЦИК СССР. – 1934.
60. Мороз М.В. Клиническая эффективность применения тауфонау больных механической желтухой: Автореф. дисс. на соиск. учен. степ. канд. мед. наук Волгоград, 1999.-20 с.
61. Мухин Ю.И. Путешествие из демократии в дерьмократию и дорога обратно. – М.: изд-во «Дар». 1993. – 288 с.
62. Нейрофизиологические и эндокринно-вегетативные корреляты экспериментального эмоционального стресса // Сб. научных трудов Харьковского мед. ин-та. ХММ. – 1988. – 690 с.
63. Неруш П.А. Влияние диазепама на некоторые показатели высшей нервной деятельности и аминокислотный состав структур мозга крыс в условиях хронического нейрогенного стресса // Физиол. журн. – 1988. – т. 34. — № 2. – с. 94-99
64. Нестурх М.Ф. Происхождение человека // Изд. «Наука» — 1970. – 440 с.
65. Нефедов Л.И. Проявление биологической активности таурина.// Изв. АН
66. Нечипоренко С.П.,Ротенберг Ю.С. Метаболизмароматических углеводородов// Итоги науки и техники.-М: ВИНИТИ-1981.-С.117-156 Беларуси. Сер. биол. наук-1992.-№ 3-4.-С.99-106.
67. Нефедов Л.И. Таурин (биохимия, фармакология и медицинское приме¬нение). — Гродно, 1999. — 140 с.
68. Нефедов Л.И., Мороз А.Р., Островский Ю.М. Статистический анализ формирования фонда свободных аминокислот печени крыс // Вопросыпитания. — 1991. — № 4.-C.37-41.
69. Новоженов Ю.И. Филетическая эволюция человека. Екатеринбург: Банк Культурной информации, 2005. – 124 с. Издание второе.
70. Новоженов Ю.И. Статус Секс и эволюция человека. Свердловск. Изд-во Ур. Унивеститета. 1991г. 160.
71. Павлов В.А. Механизмы повреждения и адаптации в организме при экспериментальном туберкулезе в условиях действия ароматических ксенобиотиков. Дисс. докт. мед. наук. – Екатеринбург. –2000. –318 с.
72. Павлов В.А., Сабадаш Е.В. Аминокислоты в механизмах резистентности к туберкулезной инфекции // Мониторинг, выявление, диагностика, лечение туберкулеза: Материалы науч.-практич. Конф.-Екатеринбург, 2004.-С.172-178.
73. Павлов В.А., Сабадаш Е.В. Аминокислоты в патогенетическом лечении
туберкулеза в современных условиях.// Материалы юбилейноц сессии.
80-летие центрального НИИ туберкулеза РАМН. 75 лет со дня
рождения академика РАМН А.Г.Хоменко-М.,2001 .-С.59-60.
74. Павлов В.А., Сабадаш Е.В. Перспективы применения аминокислот-
адаптогенов в патогенетической терапии ряда патологических
состояний организма// Пробл. Туберкулеза.-2002.-№1Л.*-С,26-28.
75. Панин Л.Е. Биохимические механизмы стресса. -Новосибирск: Наука, 1983. — 231 с.
76. Поздеев В.К. Медиаторные процессы и эпилепсия. Л.: Наука. 1983. –112 с.
77. Покровский А.А., Тутельян В.А. Лизосомы. — М.: Наука, 1976. — 382 с.
78. Поршнев Б.Ф. О начале человеческой истории. –М.: Мысль. –1974. –486 с.
79. Поршнев Б.Ф. Социальная психология и история .-М.: Наука. –1979. 231 с.
80. Придо Т. Кроманьонский человек. Пер. с англ. И. Гуровой. Ред. и предис. Ю.Г. Рычкова. М., «Мир», 1979. – 158 с.
81. Раевский К.С., Георгиев В.П. Медиаторные аминокислоты. Нейрофармакологические и нейрохимические аспекты. Совместное издание СССР-НРБ. К.: Медицина. 1986. – 240 с.
82. Робу А.И. Стресс и гипоталямические гормоны (ин-т зоологии и физиологии). Кишинев, Штиница. – 1989. – 219 с.
83. Россия между Европой и Азией: Евразийский соблазн. Антология. / Составители: Новикова Л.Н., Сиземская Я.Н. Наука. 1983. – 368 с.
84. Ростовщиков В.В. Применеие тауфона в комплексном лечении больных позднего возраста с психоорганическим синдромом. Автореф. диена соиск. учен. степ. канд. мед. наук.- Волгоград, 1999.-19 с.
85. Ростовщиков В.В. Применение тауфона в геропсихиатрической практике// Проблемы геронтологии и гериатрии: (Медицинские и социальные
аспекты): Материалы научной конференции, г. Волгоград, 1998-С.20-21.
86. Самюэль К. и др. Эволюционные подходы к связи стресса и недостаточности репродуктивной функции, в числе преждевременных родов // Междунар. медицин. журнал. – 2000. — № 2. – с. 168-171
87. Семагин В.Н., Зухарь А.В., Куликов М.А. Тип нервной системы. Стрессоустойчивость и репродукционная функция. М.: Наука. 1988. – 135 с.
88. Сент-Илер Ж. Очерки по сравнительной анатомии. – М.: Наука. – 1966.
89. Середенин С.Б. Влияние эмоционального стресса на частоту храмосомных аберраций в клетках мышей // Бюлл. экспер. биол. и медицины. – 1980. – т. 90. № 7. – с. 91-92
90. Судаков К.В. Морфофункциональные изменения нейронов мозга в условиях эмоционального стресса // Вест АМН СССР. – 1990. № 2. – С.11-13.
91. Судаков К.В. Новые акценты классической концепции стресса // Бюл. эксперим. биол. и медицины. – 1997. -№ 2. –С. 124-130.
92. Судаков К.В. Церебральные механизмы эмоционального стресса // XIX физиологического общества имени И.П. Павлова. г. Екатеринбург 19-24 сентября 2004.
93. Судаков К.В. Эмоциональный стресс как фактор эволюции // Вест. АМН СССР. – 1984. — № 6. — с. 40-45
94. Фрэзер Дж. Дж. Золотая ветвь: Исследование магии и религии: В 2 т. Т. 1: Гл. I-XXXIX // Пер. с англ. М. Рыклина. – М.: ТЕРРА-Книжный клуб, 2001. – 528 с. – (Боги и ученые).
95. Фрэзер Дж. Дж. Золотая ветвь: Исследование магии и религии: В 2 т. Т. 2: Гл. XL-LXIX // Пер. с англ. М. Рыклина; Послесл. Малиновский Б. / Пер. с англ. И. Утехина. – М.: ТЕРРА-Книжный клуб, 2001. – 496 с. – (Боги и ученые).
96. Хакам-Бей. Хаос и анархия. Революционная сотериология. М.: Гилея. – 2002. – 172 с.
97. Хочачка П., Сомеро Д.Ж. Биохимическая адаптация: пер. с англ. М.: Мир,1976.
98. Худолей В.В. Эволюция, экология и рак // Эксперим.онкология.-1993.-№ 2.- С.3-8.
99. Худолей В.В., Мизгирев И.В. Экологически опасные факторы. — СПб.: Банк Петровский, 1996. — 186 с.
100. Хьюз М. Неорганическая химия биологических процессов: пер. с англ. -М.: Мир, 1983.- 416 с.
101. Цанев Р.Г. и Марков Г.Г. Биохимия клеточного деления. – М.: Медицина. – 1964. – 120 с.
102. Чирков А.М. и др. Психофармокология эмоционального стресса у обезьян. Тбилиси: Мецниереба. – 1989. – 508 с.
103. Шабад Л.М. Методы определения и изучения бластомогенных химических веществ. — М.: Мед. , 1970. — 240 с.
104. Шапот В.С. Биохимические аспекты опухолевого роста. -М.: Мир, 1975. -304 с.
105. Шахматов И.И. отбор информативных методов оценки гомеостаза при остром эмоциональном и физическом стрессе // Клинич. лаб. диагностика. 2002. — № 9. – с. 24
106. Шейбак Л.Н., Шейбак В.М. Биологическая роль таурина у животных и человека. Обзор/ /Здравоохранение. -1996.-№2.-С.39-40.
107. Эверли Д.С., Розенфельд Р. Стресс, природа и лечение. М.: Медицина. 1985. –223 с.
108. Энгельс Ф. Происхождение семьи частной собственности и государства.- М. : Политиздат 1980, — 283 с.
109. Энциклопедический словарь. Издатели Ф.А. Брокгаузъ (Лейпциг), И.А. Ефронъ (С.-Петербург). Типо-литография И.А. Ефрона. № 6. 1896.
110. Юматов Е.А. Нейромедиаторная интеграция эмоционального возбуждения и механизмы устойчивости к стрессу // Вест. Рос. АН. –1995. -№ 11. –С.9-16.
111. Яковлев Г.М. и др. Резистентность, стресс, рауляция. Л.: Наука. Ленинград. отд. 1990. – 237 с.
112. Abdul-Chani A.S. etcol. Effect of j – acetylenic GABA and j –vinyl GABA on synaptosomal release and uptake of GABA – Brochem. Pharmacol. 1981. – v. 30. – p. 1203 – 1209
113. Hinchman Ch.A., Ballatori N. Glutathione conjugation and conversion to mercapturic acids occure as an intrahepatic process // J. Toxicol. and Environ. Health. — 1994. — V.41, N 4. — P.387-409.
114. Ho D., Fahl W.E. Modification of glutathione levels in C3H/10T1/2 cells and its relationship to benzo(a)pyrene-anti-7,8-dihydrodiol-9,10-epoxideinduced cytotoxicity // J. Biol. Chem. 1984. — V.259, N 18. — P.11231-11235.
115. Huxtable R.I. Taurine: epilepsy, intropi and eyesight. – Treuds Pharcie. Sci. – 1982. – v. 3 (1). – p. 21-25
116. Korubrast D.J., Bas J.S. Glutathione depletion by methyl chloride and association with lipid peroxidation in mice and rats // Toxicol. and Appl. Pharmacol. — 1984. — V.72, N 3. — P.388-399.
117. Lonsdale D. Free oxygen radicals and disease // Year Nutr. Med.,1986. — New Canaan, Conn., 1986. — P.85-113.
118. Luhmann N. “The Autopsies of Social systems”
119. Luisi Pier Luigi. About various Definitions of Life // Origins of Life and Evolution of the Biosphere. – 1993. – v. 28. – pp. 613-622
120. Margulis Lynn. symbiotic Planet. Basic Books. New York. 1998
121. Maturana H. “Antopoiesis and Cognition”. 1980.
122. Maturana H. Biology of Cognition Maturana H. and Varela F. 1970.
123. Maturana H., Varela F. “The Tree of Knowledge” ghambhala, Boston. 1987.
124. Maurer G.L. “Einleitung zur Geschichte der Markt – Hot – Dorf und Stadt Verfassung und der ;ffentl Gewelt”. 1854
125. Maurer G.L. “Geschichte der Markenverfassung”, 1856
126. Maurer G.L. Dorverfassung in Deutschland”. 1869
127. Oguro T., Numazava S.,Yoshida T., Biochem and Byophys. Res. Commun.-1987.-V.148, N 1.-P.422-428
128. Rassin D.K. The function of taurine in the central nervous system In: Amino acid neuro trans mitters. Ed. F. V. De Fendis, P. Mandel. New York, Raven Press, 1981, 127-134
129. Reed D.J. Glutathione: Toxicological Inplications // Annu. Rev. Pharmacol. and Toxicol. Vol. 30. — Palo Alto (Calif.), 1990. — P.603-631.
130. Schausboe A., Larsson O.M., Wood I.D. Iransport and metabolism of amino butyric aeid in neurons and glia: impliation for epilepsy // Epilepsia. 1983. – v. 24. – p. 531-538
131. Shank R.P. Apr: son significance of glutamine ajcle in neuraltissues // Life Sci. – 1981. – v. 28. p. 837-842
132. Sugiyama Y., Kaplowitz N. Binding of glutathione by rat liver cytosol // Pharmacology. — 1984. — V.28, N 2. — P.61-66.
133. Varela F. “Neurophenomenology // Journal of Consciousness Studies. – 1996. – Vol. 3. no. 4, pp. 330-349
134. Varela F. Present – Time Consciousness // Journal of Consciousness Studies. – 1999. – Vol. 6. no. 2-3, pp. 111-140
135. Varela F. Resonant Cell Assambles // Biological Research, 1995. – Vol. 28. – pp. 81-95
136. Videla L.A. Chemically induced antioxidant sensitive respiration. Relation to glutathione content and lipid peroxidation in the perfused rat liver // FEBS Lett. — 1984. — V.178, N 1. — P.119-122.
137. Whight C.E., Tallan H.H., Lin Y.Y., Gauff C.E. Taurine: Biological update // Annu. Rev. Biochem. — Vol. 55. — Palo Alto (Calif.). — 1986. — P.427-453.
СПИСОК СОКРАЩЕНИЙ
КХА – катехоламины
ГК – глюкокортикоиды
БАВ – биологически-активные вещества
ПАУ – полициклические ароматические углеводороды
ОВП – окислительно-восстановительные процессы
ОСФ – оксигензы смешанных функций
ГМФШ – гиксозомонофосфатный шунт (пентозный цикл)
ФАФС – фосфоаденозин фосфосульфат
УДФГК – уридиндифосфоглюкороновая кислота
ЗАМ – защитно-адаптивные механизмы
АФК – активные формы кислорода
ПОЛ – перекисное окисление липидов
ГГ – глюконеогенез
Г-SH – глутатион восстановленный
Г-SS-Г – глутатион окисленный
ЦНС – центральная нервная система
НЭС – нейроэндокринная система
АТФ – аденозинтрифосфат
АДФ – аденозиндифосфат
АМФ – аденозинмонофосфат
ЦАМФ – цеклический аденазинмонофосфат
ЦГМФ – цеклический гуанозинмонофосфат
ГЭБ – гематоэнцефалитический барьер
ЭС – эмоциональный стресс
АК – аскорбиновая кислота
СТ – соединительная ткань
ЦТК – цикл трикарбоновых кислот (цикл Кребса)
РНК – рибонуклеиновая кислота
ДЦ – дыхательная цепь
Э-М-К – Эмбдена-Мейергофа-Кребса (путь метаболизирования углеводов)
ГОМК – гаммаоксимасляная кислота
ГАМК – гаммааминомасляная кислота
НАДН2 – никотинамидадениндинуклиотид восстановленный
НАДФН2 – никотинамидадениндинуклиотидфосфат восстановленный
ЛПОНП – липопротеиды очень низкой плотности
ЛПНП – липопротеиды низкой плотности
ЛПВП – липопротеиды высокой плотности
СЖК – свободные жирные кислоты